

β-1,6-Glucan plays a central role in the structure and remodeling of the bilaminate fungal cell wall
- PMID: 39636210
- PMCID: PMC11620752
- DOI: 10.7554/eLife.100569
β-1,6-Glucan plays a central role in the structure and remodeling of the bilaminate fungal cell wall
Abstract
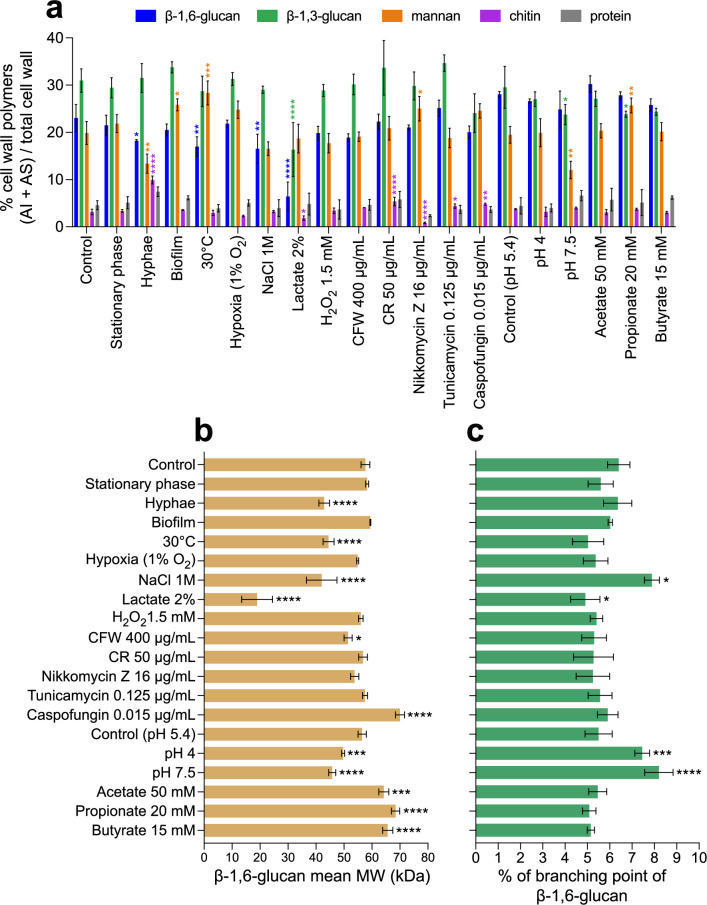
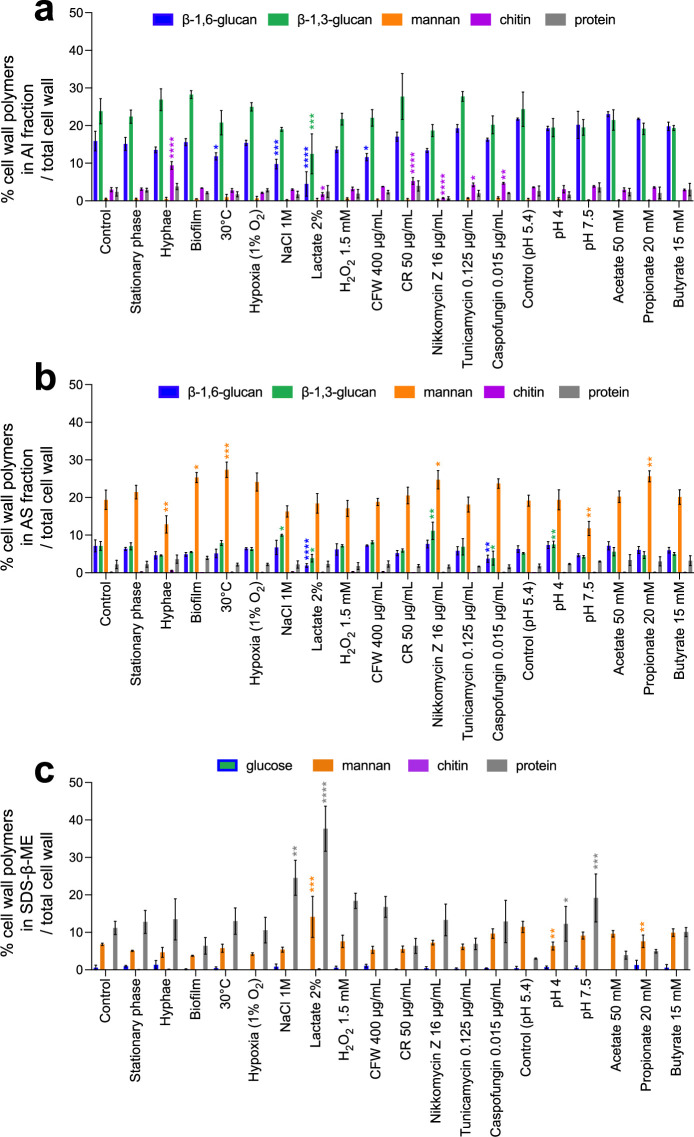
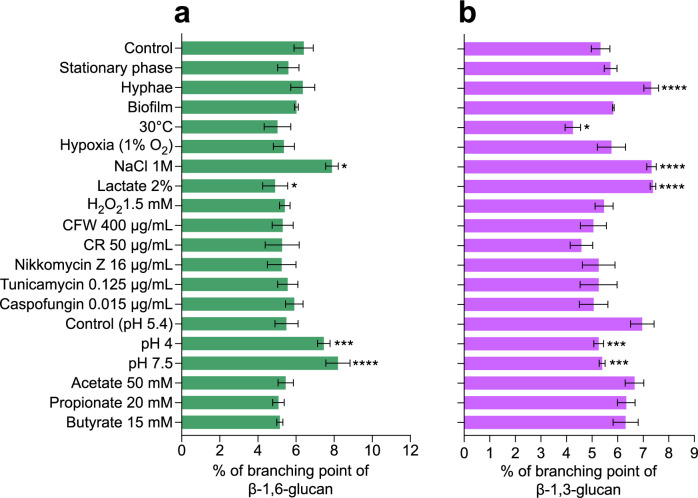
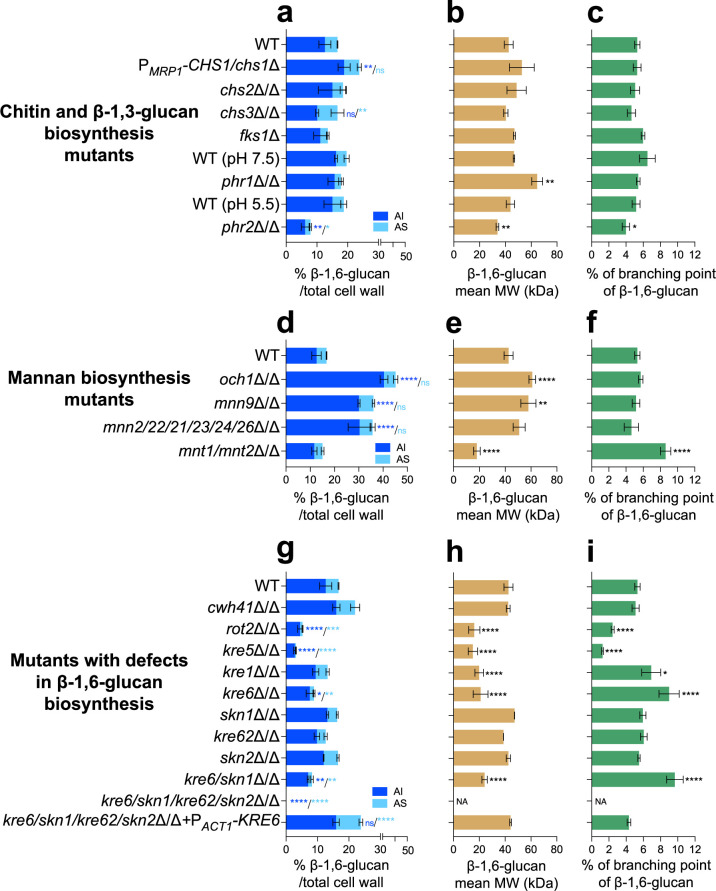
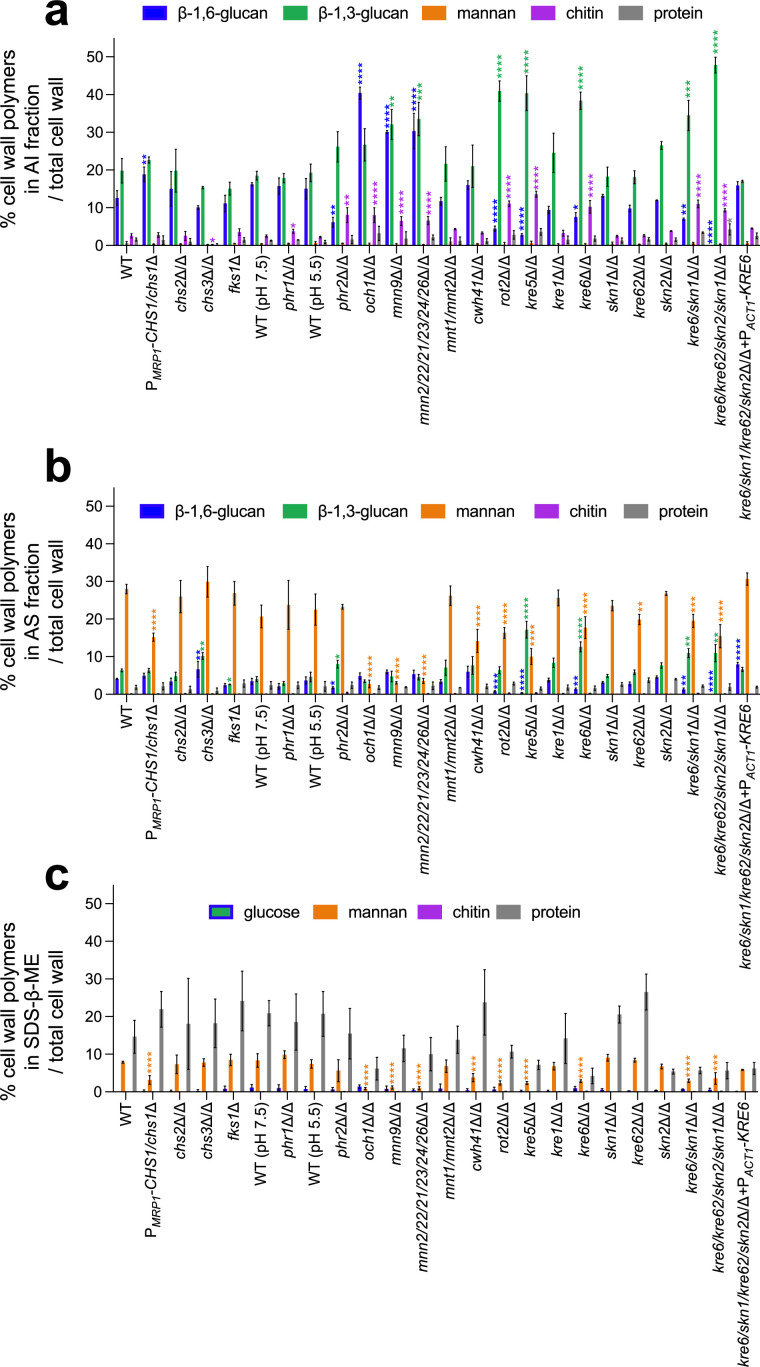
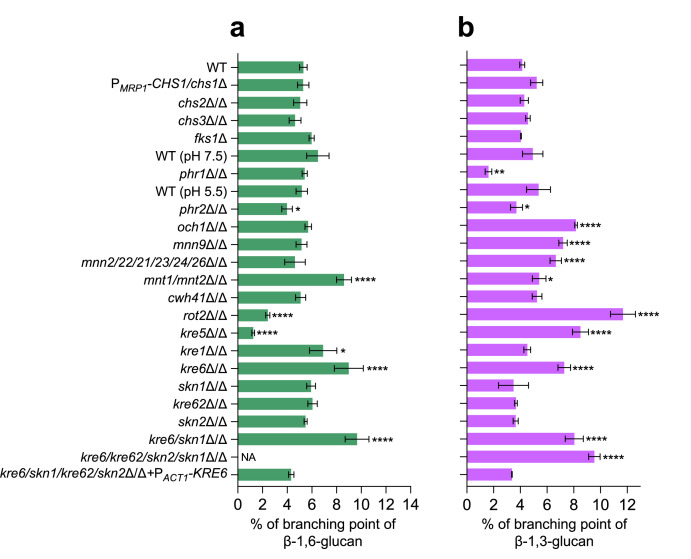
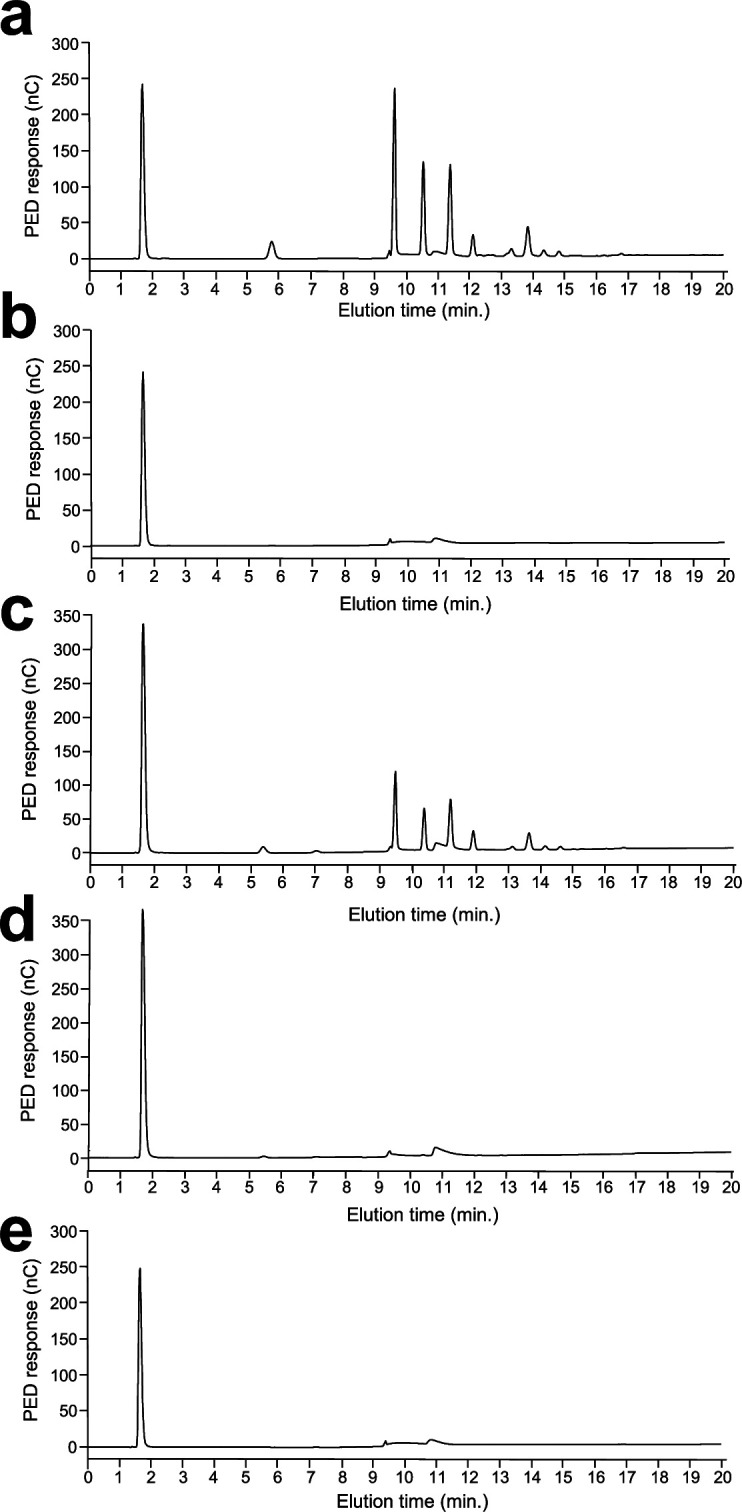
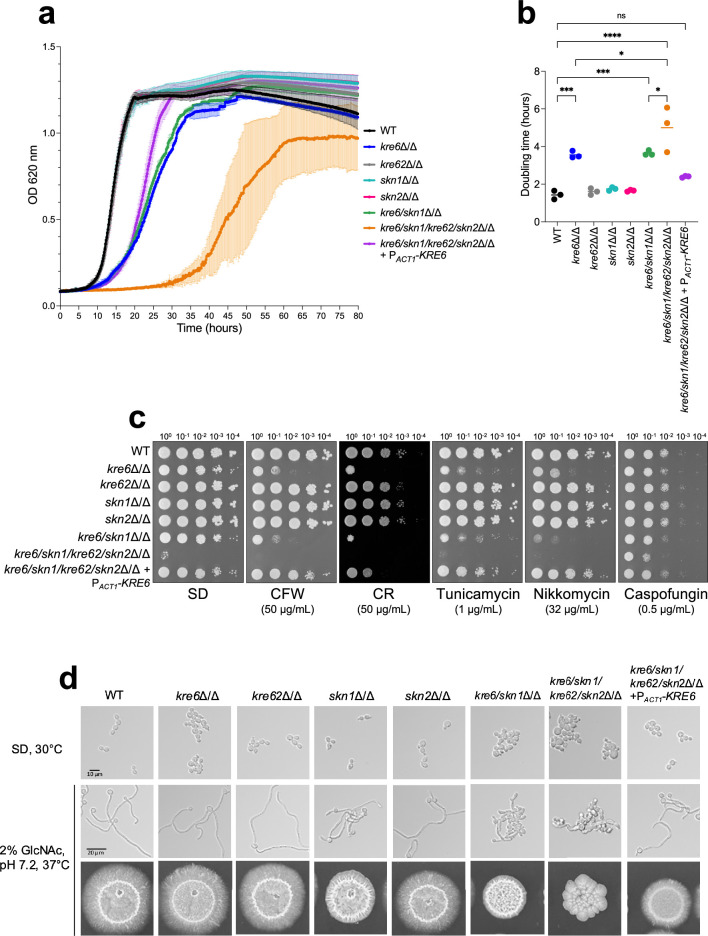
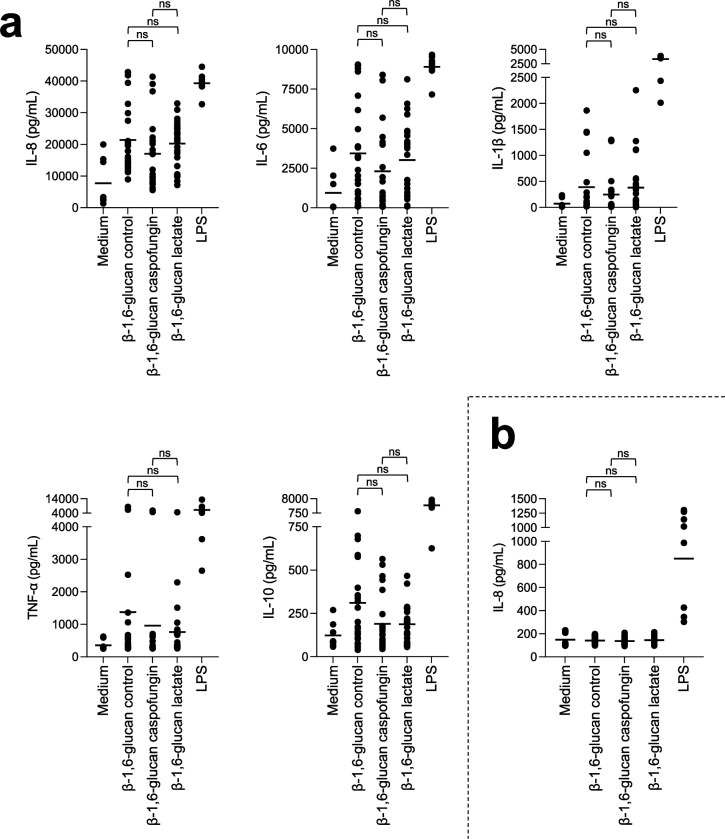
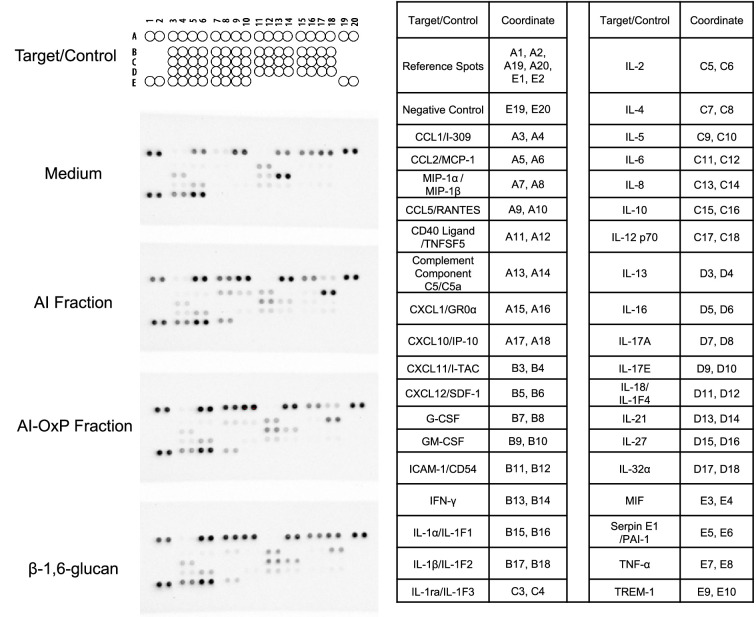
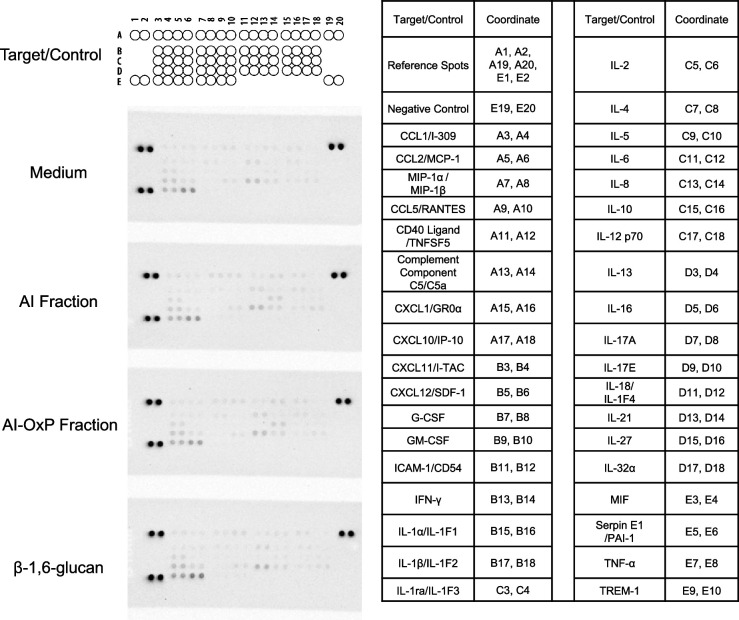
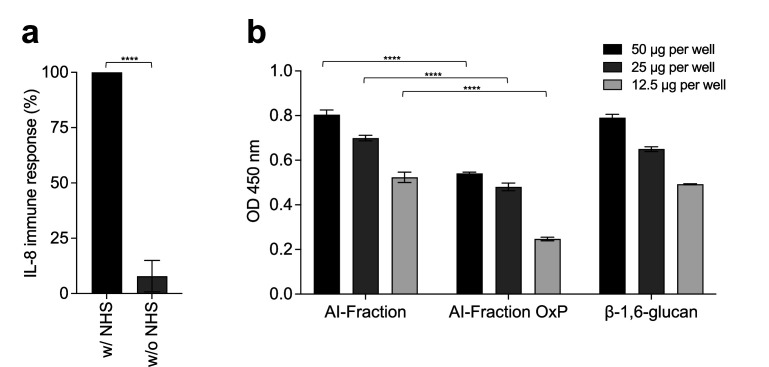
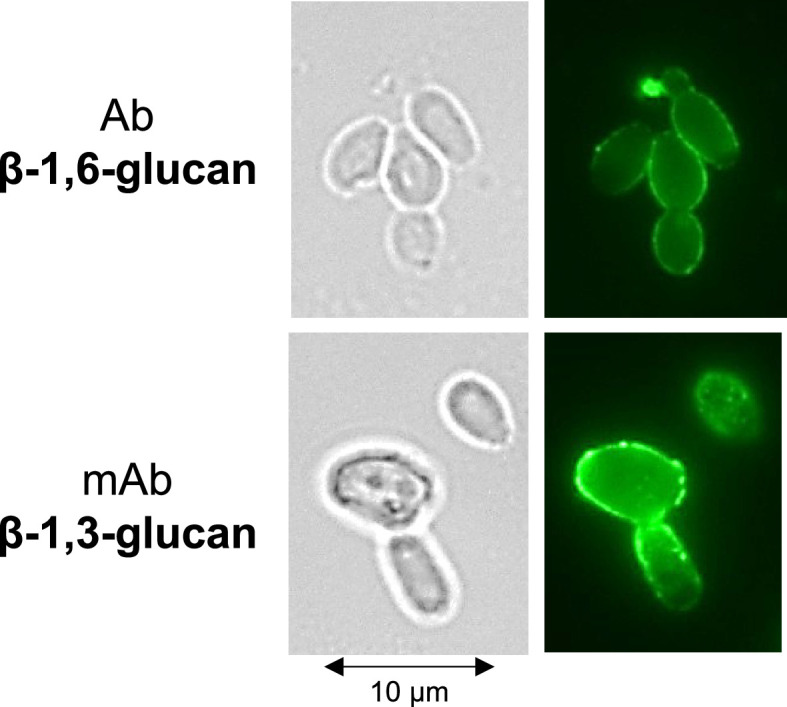
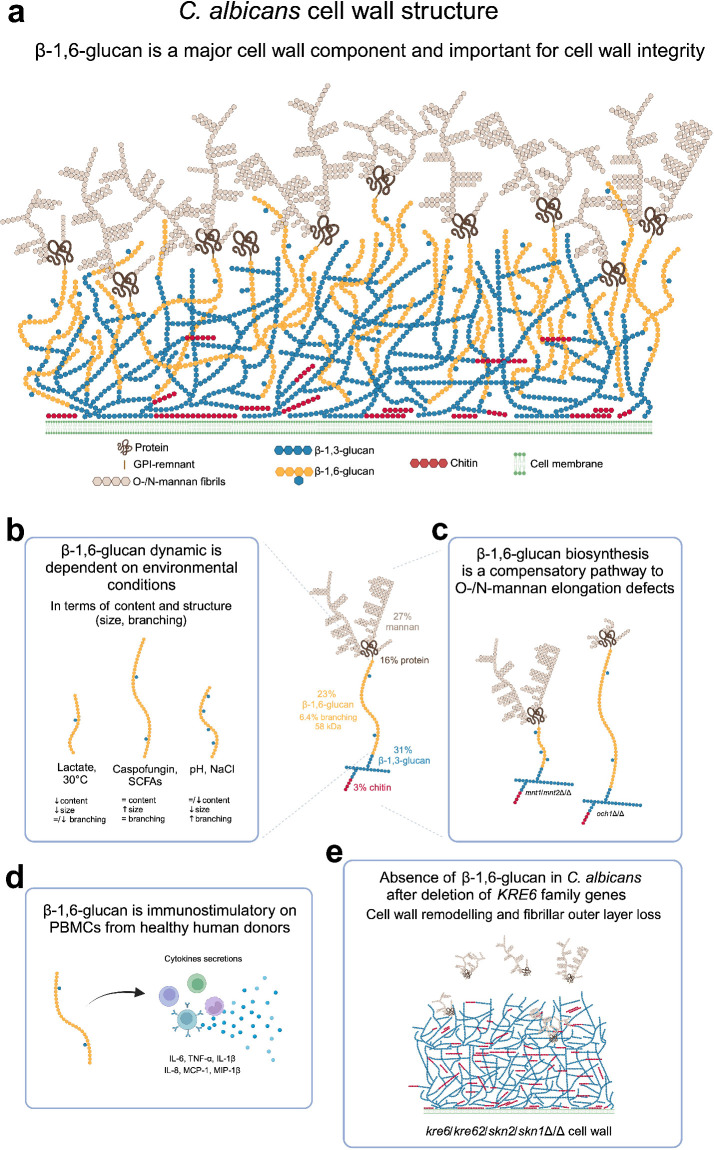
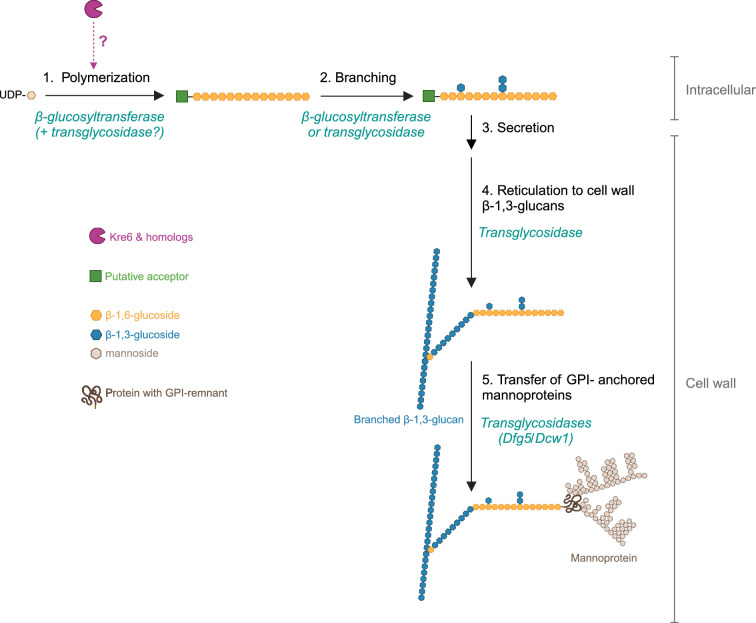
The cell wall of human fungal pathogens plays critical roles as an architectural scaffold and as a target and modulator of the host immune response. Although the cell wall of the pathogenic yeast Candida albicans is intensively studied, one of the major fibrillar components in its cell wall, β-1,6-glucan, has been largely neglected. Here, we show that β-1,6-glucan is essential for bilayered cell wall organization, cell wall integrity, and filamentous growth. For the first time, we show that β-1,6-glucan production compensates the defect in mannan elongation in the outer layer of the cell wall. In addition, β-1,6-glucan dynamics are also coordinated by host environmental stimuli and stresses with wall remodeling, where the regulation of β-1,6-glucan structure and chain length is a crucial process. As we point out that β-1,6-glucan is exposed at the yeast surface and modulate immune response, β-1,6-glucan must be considered a key factor in host-pathogen interactions.
Keywords: Candida albicans; KRE6; biochemistry; cell wall; chemical biology; immune response; infectious disease; microbiology; β-1,6-Glucan.
© 2024, Bekirian et al.
Conflict of interest statement
CB, IV, SB, CS, JG, MC, TM, NG, VA, Cd, TF No competing interests declared
Figures
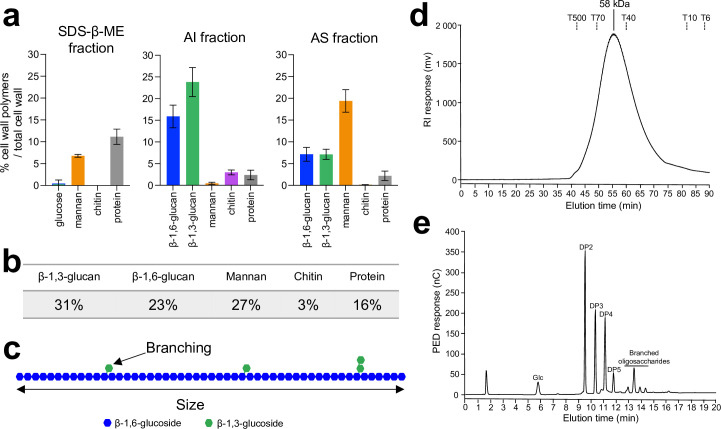
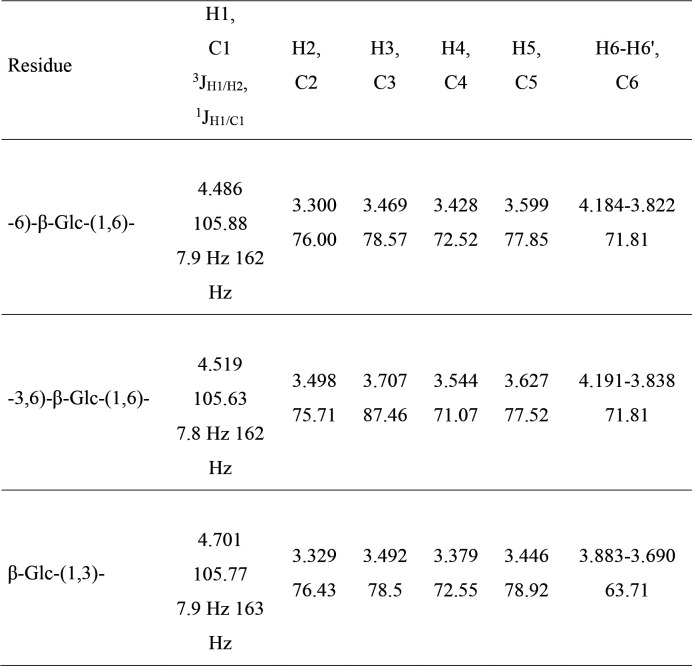
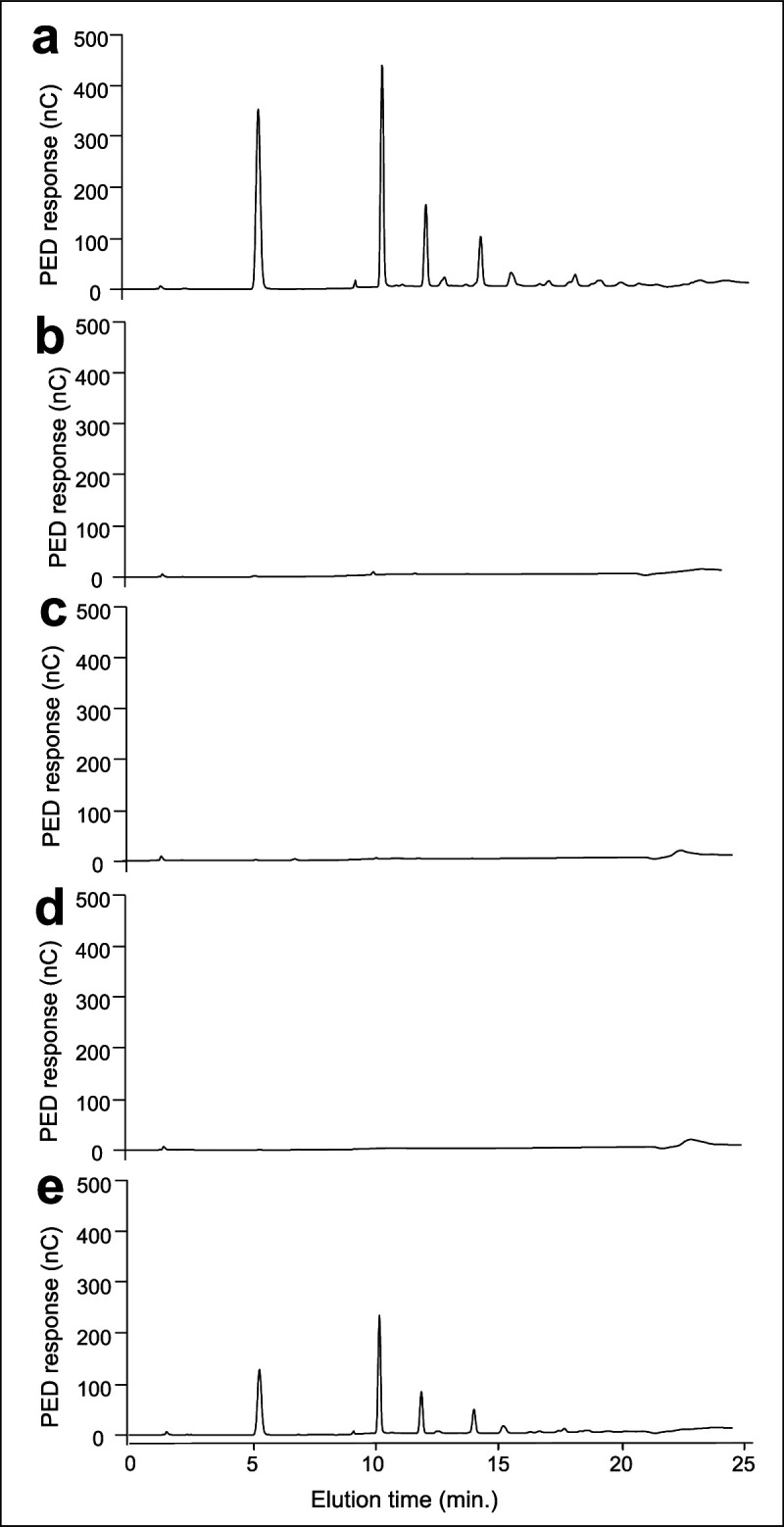
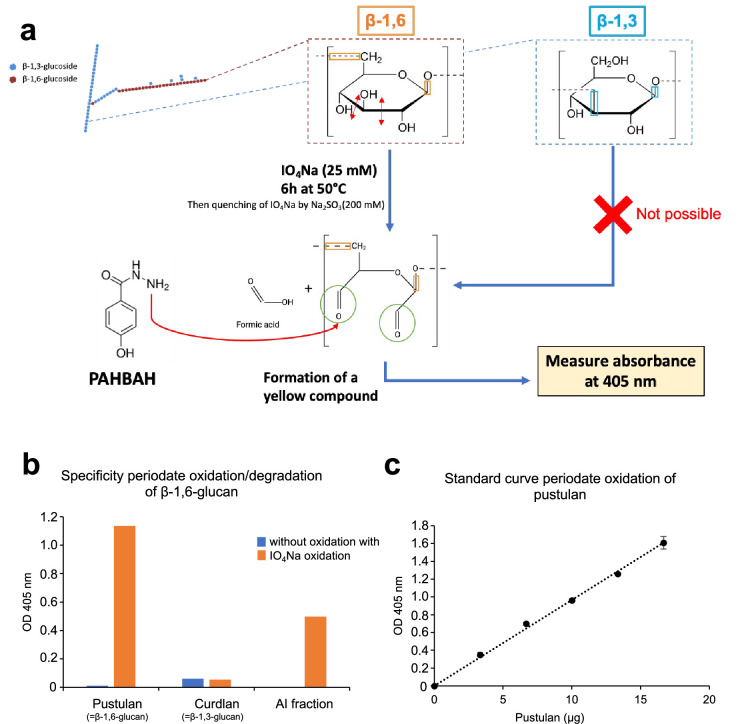
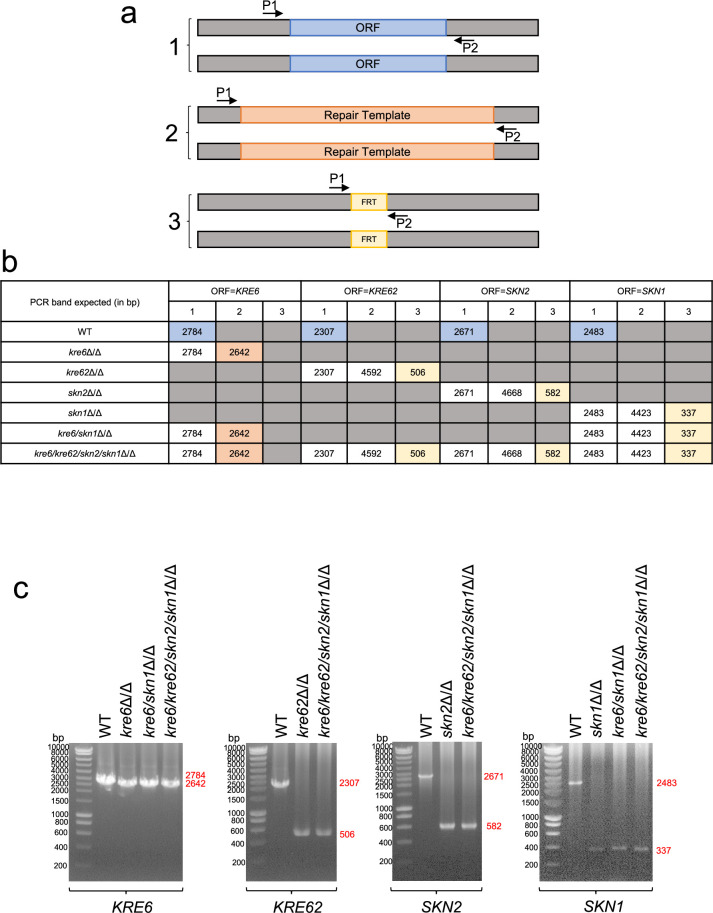
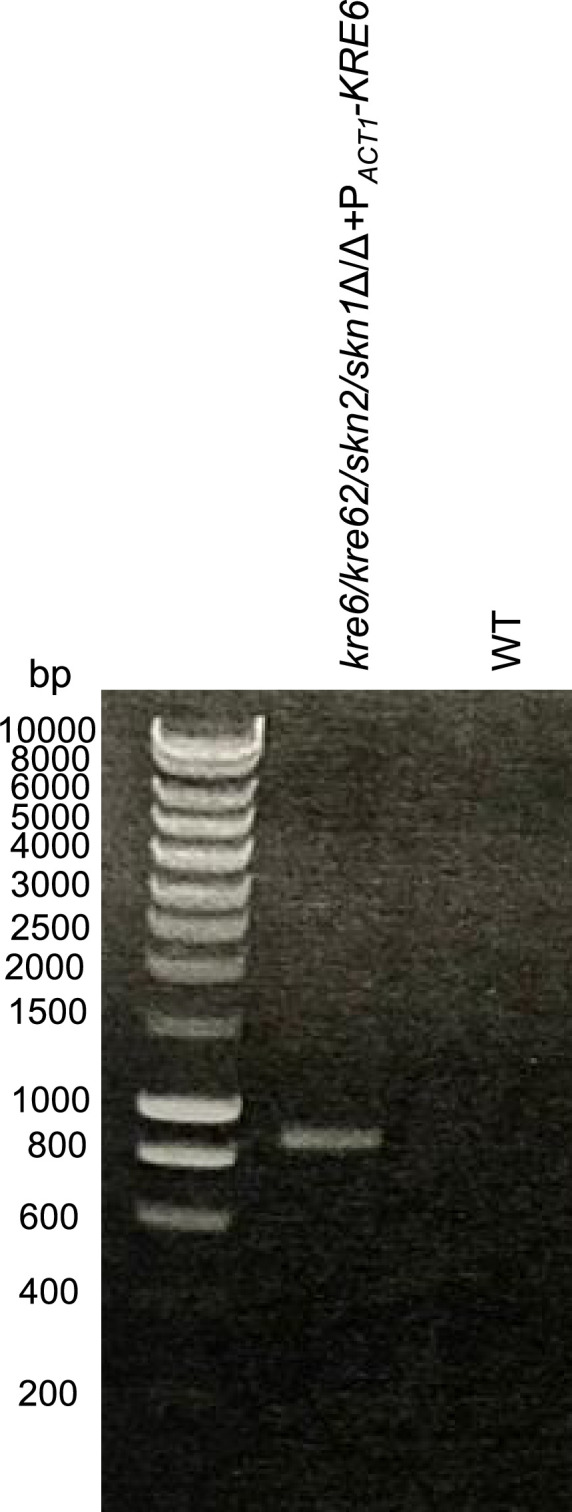
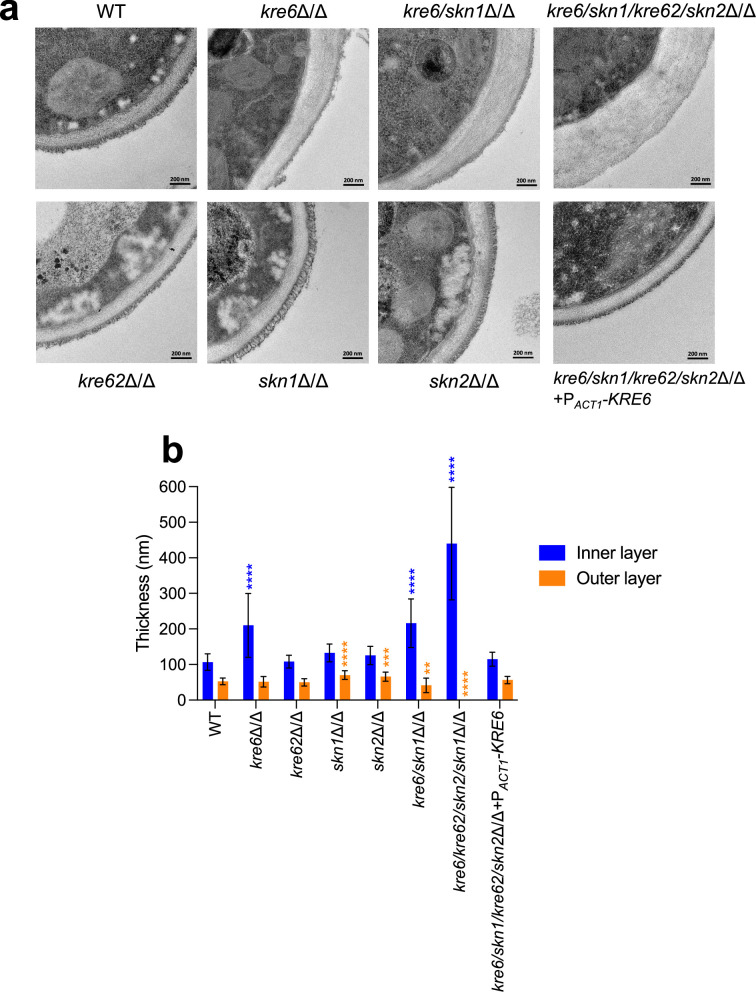
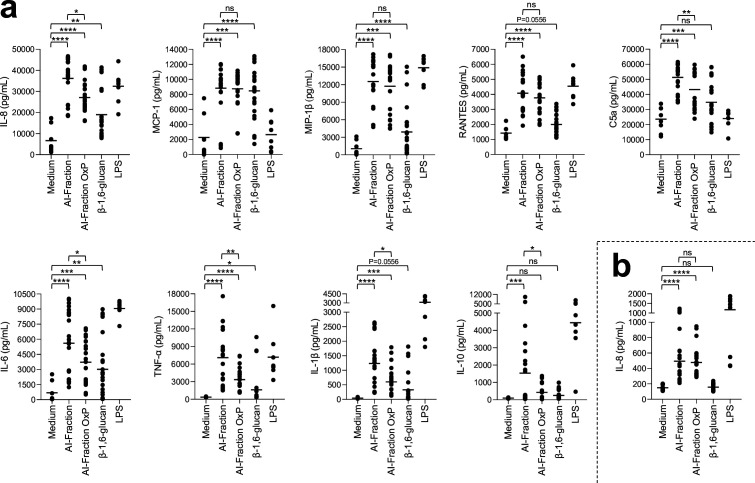
Update of
References
-
- Aimanianda V, Simenel C, Garnaud C, Clavaud C, Tada R, Barbin L, Mouyna I, Heddergott C, Popolo L, Ohya Y, Delepierre M, Latge JP. The dual activity responsible for the elongation and branching of β-(1,3)-Glucan in the fungal cell wall. mBio. 2017;8:e00619. doi: 10.1128/mBio.00619-17. - DOI - PMC - PubMed
-
- Ancian B, Bourgeois I, Dauphin JF, Shaw AA. Artifact-free pure absorption PFG-Enhanced DQF-COSY spectra including a gradient pulse in the evolution period. Journal of Magnetic Resonance. 1997;125:348–354. doi: 10.1006/jmre.1997.1112. - DOI
-
- Ballou ER, Avelar GM, Childers DS, Mackie J, Bain JM, Wagener J, Kastora SL, Panea MD, Hardison SE, Walker LA, Erwig LP, Munro CA, Gow NAR, Brown GD, MacCallum DM, Brown AJP. Lactate signalling regulates fungal β-glucan masking and immune evasion. Nature Microbiology. 2016;2:16238. doi: 10.1038/nmicrobiol.2016.238. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 10.35802/200208/WT_/Wellcome Trust/United Kingdom
- MR/N006364/2/MRC Centre for Medical Mycology
- ANR-21-CE17-0032-01/Agence Nationale de la Recherche
- SESAME 2014 NMRCHR grant no 4014526/Region Ile de France
- MR/M026663/2/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- ANR-10-LABX-62-IBEID/Laboratoire d'Excellence Integrative Biology of Emerging Infectious Diseases
- 10.35802/224323/WT_/Wellcome Trust/United Kingdom
- 10.35802/215599/WT_/Wellcome Trust/United Kingdom
- 10.35802/101873/WT_/Wellcome Trust/United Kingdom
- ANR-10-INSB-04-01/French Government Programme Investissements d'Avenir France BioImaging
LinkOut - more resources
Full Text Sources
Research Materials
