

The Greatwall-Endosulfine-PP2A/B55 pathway regulates entry into quiescence by enhancing translation of Elongator-tunable transcripts
- PMID: 39638797
- PMCID: PMC11621810
- DOI: 10.1038/s41467-024-55004-4
The Greatwall-Endosulfine-PP2A/B55 pathway regulates entry into quiescence by enhancing translation of Elongator-tunable transcripts
Abstract
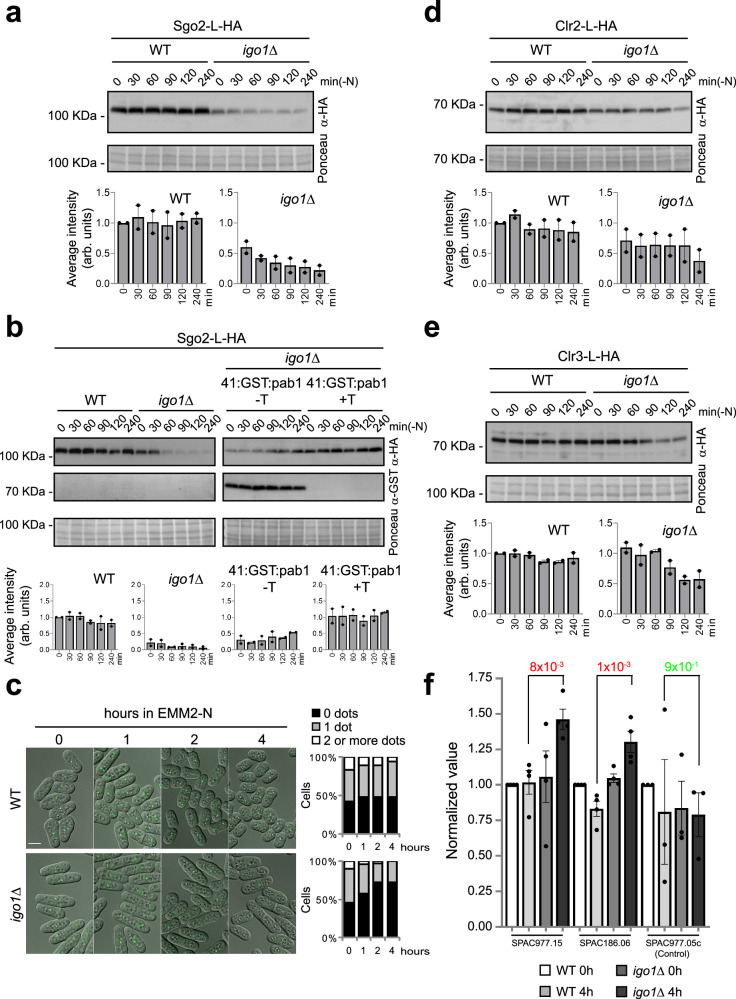
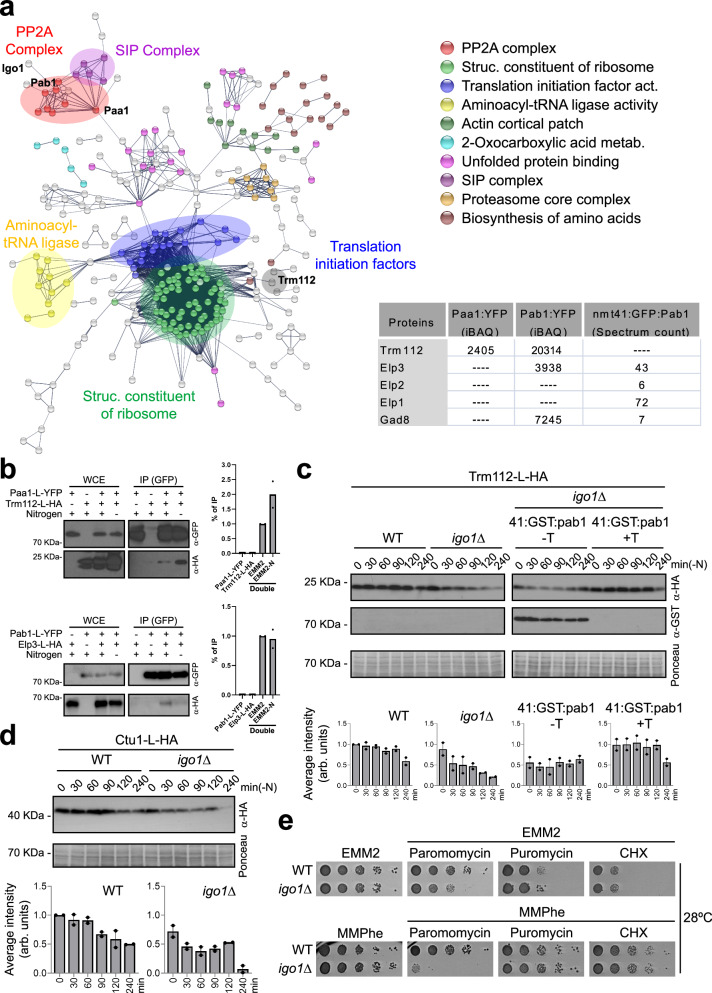
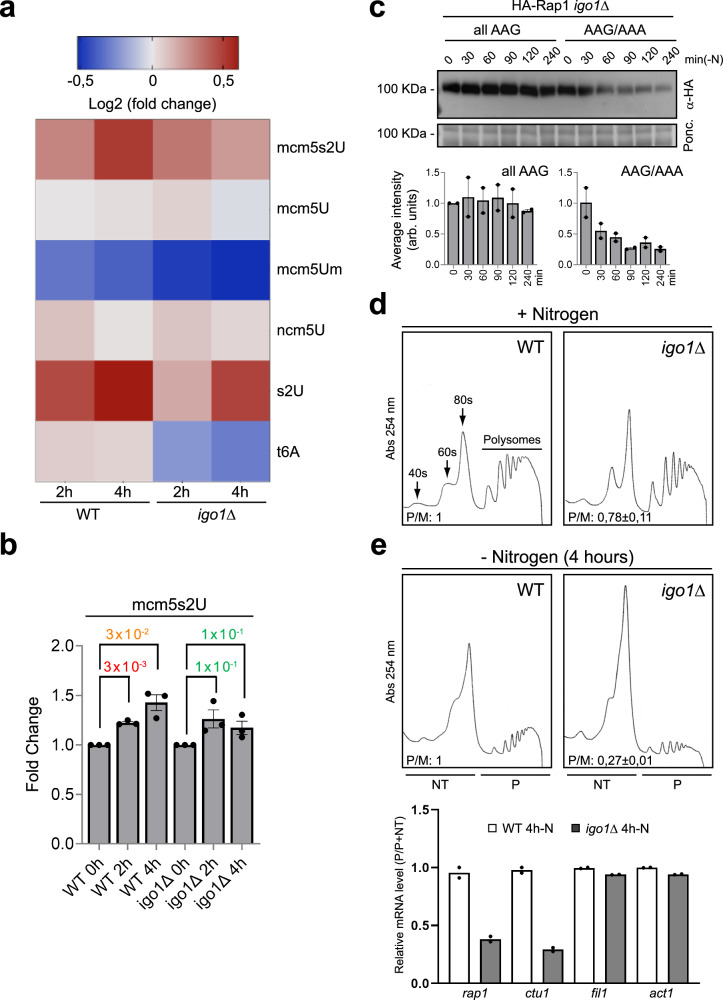
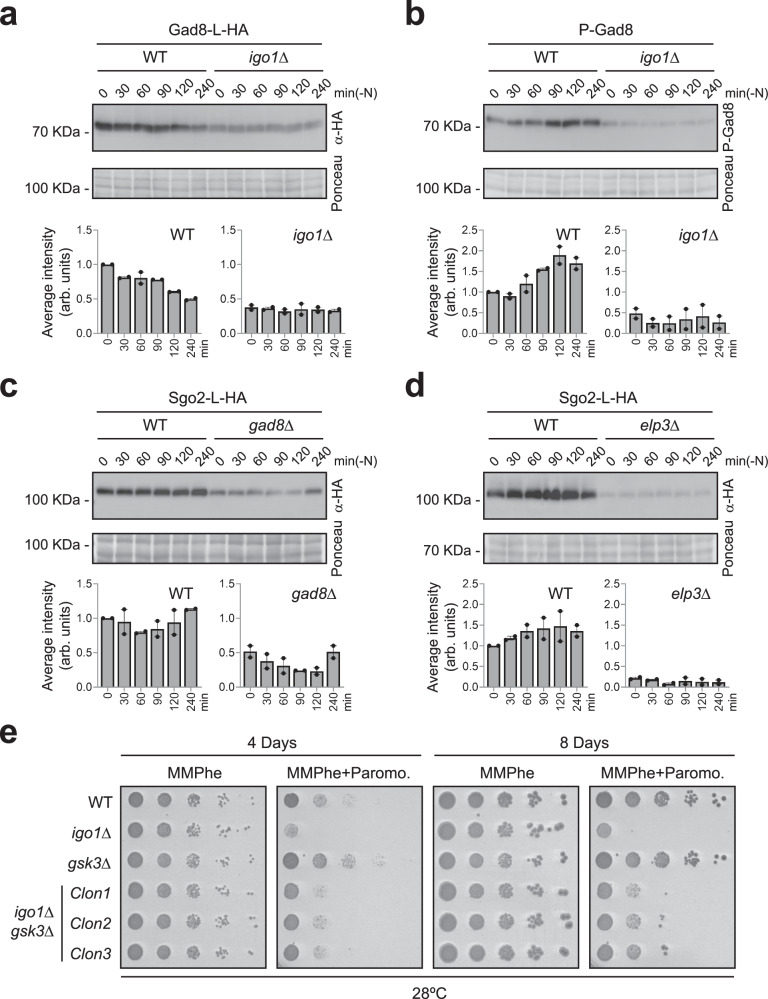
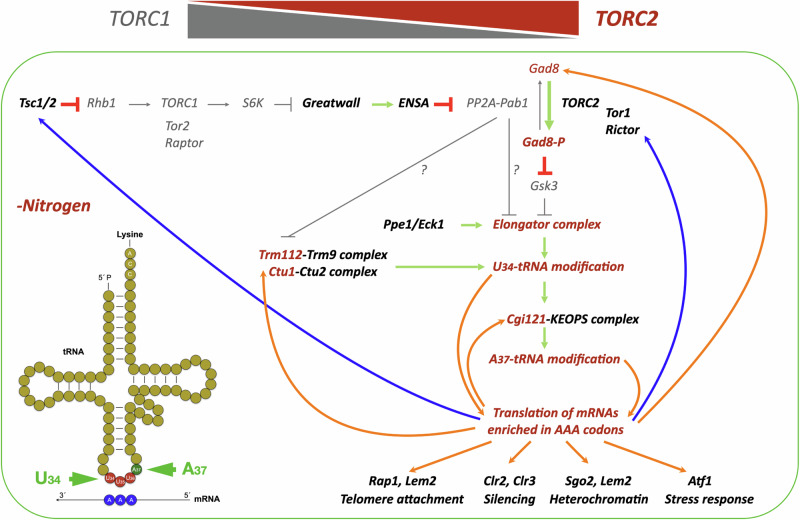
Quiescent cells require a continuous supply of proteins to maintain protein homeostasis. In fission yeast, entry into quiescence is triggered by nitrogen stress, leading to the inactivation of TORC1 and the activation of TORC2. In this study, we demonstrate that the Greatwall-Endosulfine-PPA/B55 pathway connects the downregulation of TORC1 with the upregulation of TORC2, resulting in the activation of Elongator-dependent tRNA modifications crucial for sustaining the translation programme during entry into quiescence. This mechanism promotes U34 and A37 tRNA modifications at the anticodon stem loop, enhancing translation efficiency and fidelity of mRNAs enriched for AAA versus AAG lysine codons. Notably, several of these mRNAs encode TORC1 inhibitors, TORC2 activators, tRNA modifiers, and proteins necessary for telomeric and subtelomeric functions. Therefore, we propose a mechanism by which cells respond to nitrogen stress at the level of translation, involving a coordinated interplay between tRNA epitranscriptome and biased codon usage.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


Update of
-
The Greatwall-Endosulfine-PP2A/B55 pathway controls entry into quiescence by promoting translation of Elongator-tuneable transcripts.Res Sq [Preprint]. 2023 Dec 5:rs.3.rs-3616701. doi: 10.21203/rs.3.rs-3616701/v1. Res Sq. 2023. Update in: Nat Commun. 2024 Dec 5;15(1):10603. doi: 10.1038/s41467-024-55004-4. PMID: 38105947 Free PMC article. Updated. Preprint.
References
-
- Gonzalez, A., Hall, M. N., Lin, S. C. & Hardie, D. G. AMPK and TOR: the Yin and Yang of cellular nutrient sensing and growth control. Cell Metab.31, 472–492 (2020). - PubMed
-
- Bontron, S. et al. Yeast endosulfines control entry into quiescence and chronological life span by inhibiting protein phosphatase 2A. Cell Rep.3, 16–22 (2013). - PubMed
-
- Chica, N. et al. Nutritional control of cell size by the Greatwall-Endosulfine-PP2A.B55 pathway. Curr. Biol.26, 319–330 (2016). - PubMed
-
- Aono, S., Haruna, Y., Watanabe, Y. H., Mochida, S. & Takeda, K. The fission yeast Greatwall-Endosulfine pathway is required for proper quiescence/G(0) phase entry and maintenance. Genes Cells24, 172–186 (2019). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
