

In vivo affinity maturation of the CD4 domains of an HIV-1-entry inhibitor
- PMID: 39638875
- PMCID: PMC12067531
- DOI: 10.1038/s41551-024-01289-1
In vivo affinity maturation of the CD4 domains of an HIV-1-entry inhibitor
Abstract
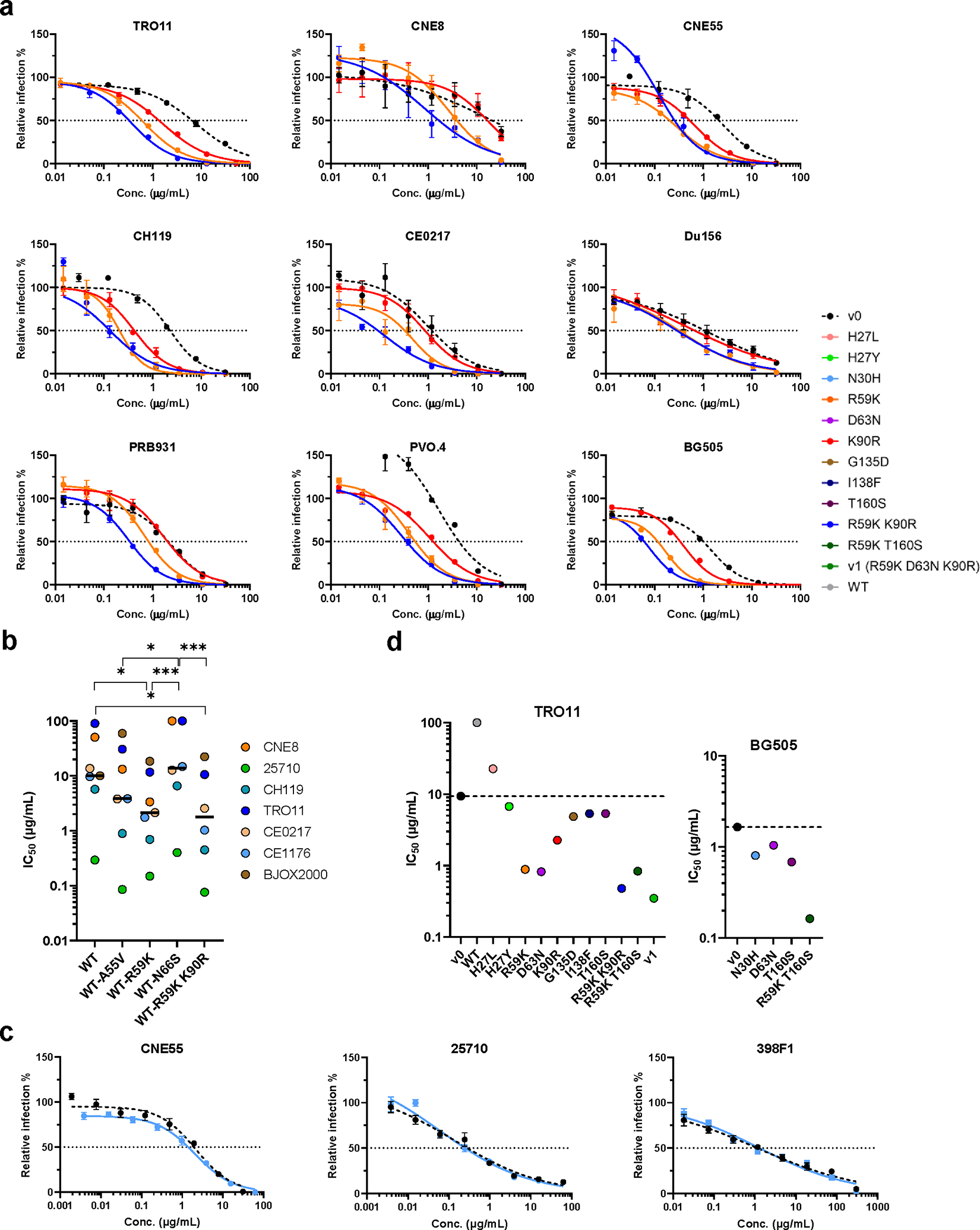
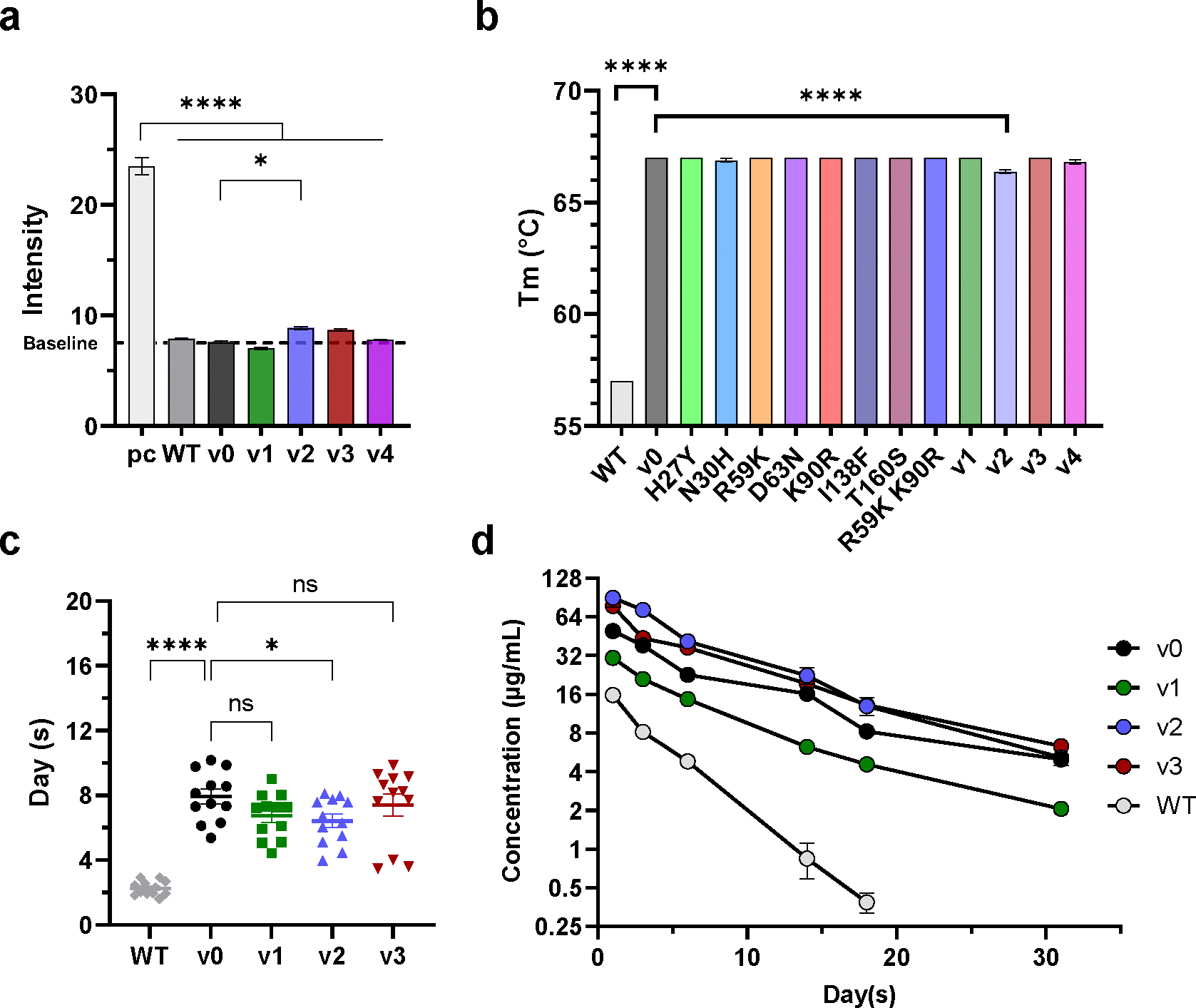
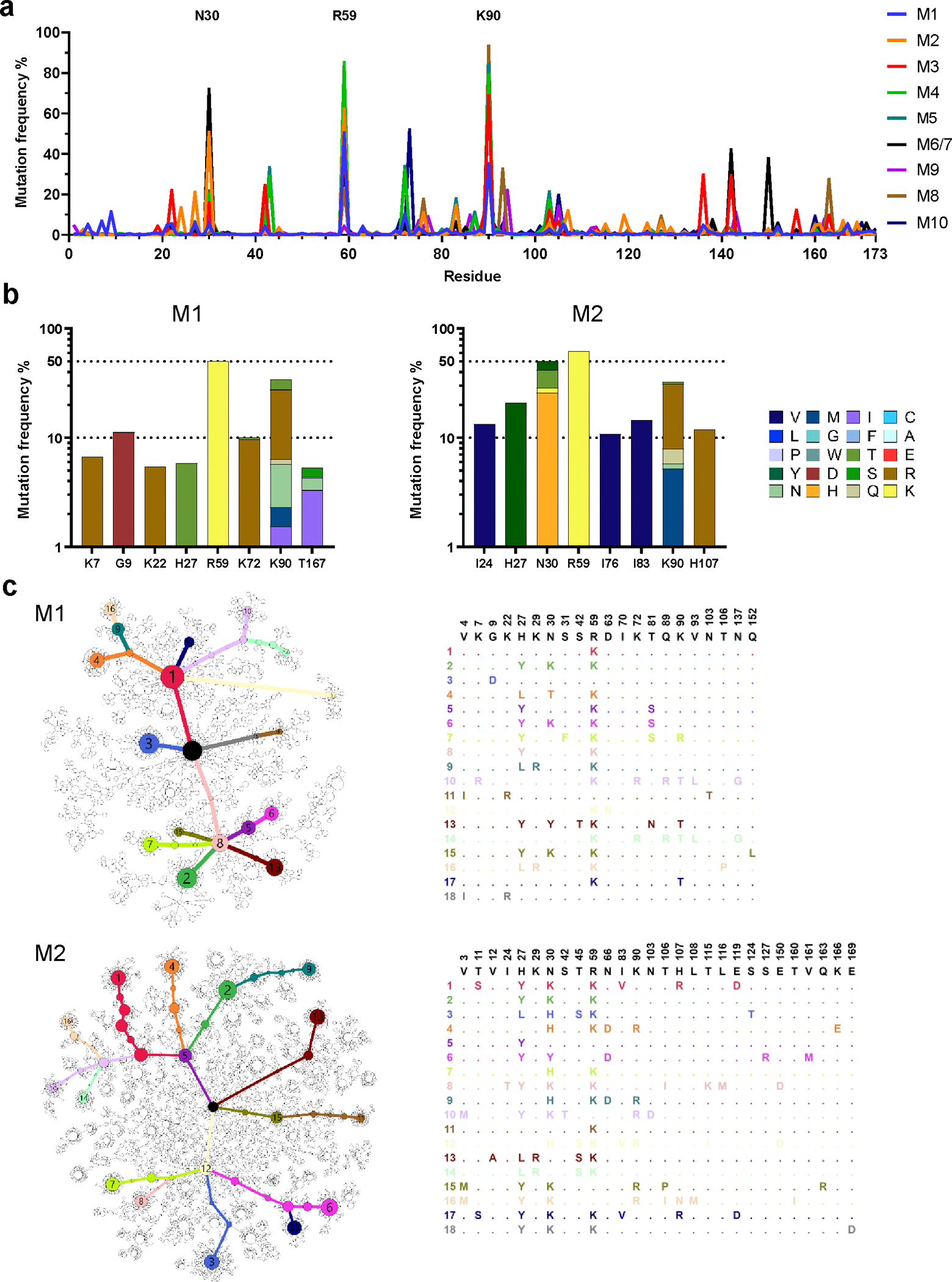
Human proteins repurposed as biologics for clinical use have been engineered through in vitro techniques that improve the affinity of the biologics for their ligands. However, the techniques do not select against properties, such as protease sensitivity or self-reactivity, that impair the biologics' clinical efficacy. Here we show that the B-cell receptors of primary murine B cells can be engineered to affinity mature in vivo the human CD4 domains of the HIV-1-entry inhibitor CD4 immunoadhesin (CD4-Ig). Specifically, we introduced genes encoding the CD4 domains 1 and 2 (D1D2) of a half-life-enhanced form of CD4-Ig (CD4-Ig-v0) into the heavy-chain loci of murine B cells and adoptively transferred these cells into wild-type mice. After immunization, the B cells proliferated, class switched, affinity matured and produced D1D2-presenting antibodies. Somatic hypermutations in the D1D2-encoding region of the engrafted cells improved the binding affinity of CD4-Ig-v0 for the HIV-1 envelope glycoprotein and the inhibitor's ability to neutralize a panel of HIV-1 isolates without impairing its pharmacokinetic properties. In vivo affinity maturation of non-antibody protein biologics may guide the development of more effective therapeutics.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: A.P., W.H., T.O., Y.Y. and M.F. are inventors of a patent describing the in vivo affinity maturation of antibodies and biologics. C.C.B., M.D.A. and M.F. have equity stakes in Emmune, Inc., which developed CD4-Ig-v0. The other authors declare no competing interests.
Figures














Update of
-
In vivo affinity maturation of the HIV-1 Env-binding domain of CD4.bioRxiv [Preprint]. 2024 Feb 5:2024.02.03.578630. doi: 10.1101/2024.02.03.578630. bioRxiv. 2024. Update in: Nat Biomed Eng. 2024 Dec;8(12):1715-1729. doi: 10.1038/s41551-024-01289-1. PMID: 38370774 Free PMC article. Updated. Preprint.
-
In vivo affinity maturation of the HIV-1 Env-binding domain of CD4.Res Sq [Preprint]. 2024 Feb 9:rs.3.rs-3922904. doi: 10.21203/rs.3.rs-3922904/v1. Res Sq. 2024. Update in: Nat Biomed Eng. 2024 Dec;8(12):1715-1729. doi: 10.1038/s41551-024-01289-1. PMID: 38405717 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- R01 AI174277/AI/NIAID NIH HHS/United States
- AI091476/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R01 AI154989/AI/NIAID NIH HHS/United States
- R01 DA056771/DA/NIDA NIH HHS/United States
- R21 AI152836/AI/NIAID NIH HHS/United States
- U19 AI149646/AI/NIAID NIH HHS/United States
- AI152836/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- AI149646/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R01 AI174270/AI/NIAID NIH HHS/United States
- DA056771/U.S. Department of Health & Human Services | NIH | National Institute on Drug Abuse (NIDA)
- R37 AI091476/AI/NIAID NIH HHS/United States
- UM1 AI126623/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
