

Natural Killer Cell-Mediated Cytotoxicity Shapes the Clonal Evolution of B-cell Leukemia
- PMID: 39642167
- PMCID: PMC7617306
- DOI: 10.1158/2326-6066.CIR-24-0189
Natural Killer Cell-Mediated Cytotoxicity Shapes the Clonal Evolution of B-cell Leukemia
Abstract
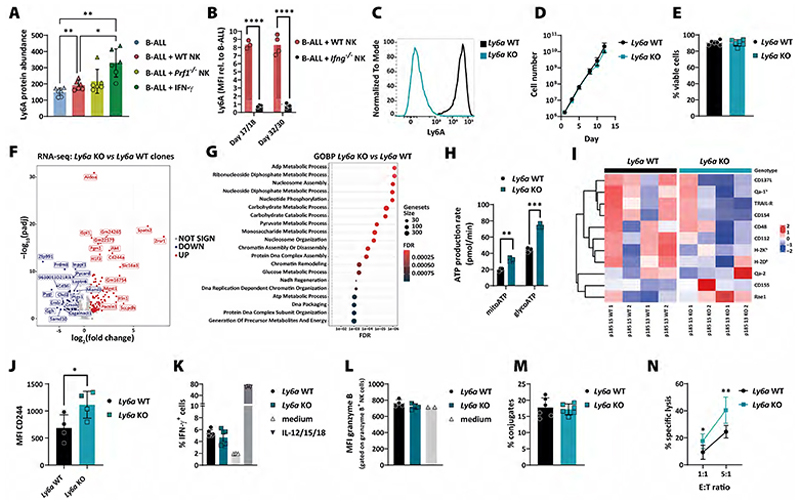
The term cancer immunoediting describes the dual role by which the immune system can suppress and promote tumor growth and is divided into three phases: elimination, equilibrium, and escape. The role of NK cells has mainly been attributed to the elimination phase. Here, we show that NK cells play a role in all three phases of cancer immunoediting. Extended co-culturing of DNA-barcoded mouse BCR/ABLp185+ B-cell acute lymphoblastic leukemia (B-ALL) cells with NK cells allowed for a quantitative measure of NK cell-mediated immunoediting. Although most tumor cell clones were efficiently eliminated by NK cells, a certain fraction of tumor cells harbored an intrinsic primary resistance. Furthermore, DNA barcoding revealed tumor cell clones with secondary resistance, which stochastically acquired resistance to NK cells. NK cell-mediated cytotoxicity put a selective pressure on B-ALL cells, which led to an outgrowth of primary and secondary resistant tumor cell clones, which were characterized by an IFNγ signature. Besides well-known regulators of immune evasion, our analysis of NK cell-resistant tumor cells revealed the upregulation of genes, including lymphocyte antigen 6 complex, locus A (Ly6a), which we found to promote leukemic cell resistance to NK cells. Translation of our findings to the human system showed that high expression of LY6E on tumor cells impaired their physical interaction with NK cells and led to worse prognosis in patients with leukemia. Our results demonstrate that tumor cells are actively edited by NK cells during the equilibrium phase and use different avenues to escape NK cell-mediated eradication.
©2024 American Association for Cancer Research.
Conflict of interest statement
Figures





References
-
- Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD. Nat Immunol. Vol. 3. Nature Publishing Group; 2002. Cancer immunoediting: from immunosurveillance to tumor escape; pp. 991–8. - PubMed
-
- Wolf NK, Kissiov DU, Raulet DH. Nat Rev Immunol. Vol. 23. Nature Publishing Group; 2023. Roles of natural killer cells in immunity to cancer, and applications to immunotherapy; pp. 90–105. - PubMed
-
- Metkar SS, Wang B, Aguilar-Santelises M, Raja SM, Uhlin-Hansen L, Podack E, et al. Immunity. Vol. 16. Elsevier; 2002. Cytotoxic Cell Granule-Mediated Apoptosis: Perforin Delivers Granzyme B-Serglycin Complexes into Target Cells without Plasma Membrane Pore Formation; pp. 417–28. - PubMed
-
- Prager I, Watzl C. Mechanisms of natural killer cell-mediated cellular cytotoxicity. J Leukoc Biol. 2019;105:1319–29. - PubMed
MeSH terms
Grants and funding
- 20-17258/Alex's Lemonade Stand Foundation for Childhood Cancer (ALSF)
- 10.55776/P32001/Austrian Science Fund (FWF)
- Fellinger Krebsforschung
- TAI 454/FWF_/Austrian Science Fund FWF/Austria
- TAI 732/FWF_/Austrian Science Fund FWF/Austria
- I 4649/FWF_/Austrian Science Fund FWF/Austria
- Dr. Mildred Scheel Stiftung für Krebsforschung
- 10.55776/TAI454/Austrian Science Fund (FWF)
- P 32001/FWF_/Austrian Science Fund FWF/Austria
- P 34832/FWF_/Austrian Science Fund FWF/Austria
- WKP 132/FWF_/Austrian Science Fund FWF/Austria
- P 31563/FWF_/Austrian Science Fund FWF/Austria
- 10.55776/P31563/Austrian Science Fund (FWF)
- St. Anna Kinderkrebsforschung
- #25905/Österreichischen Akademie der Wissenschaften (ÖAW)
LinkOut - more resources
Full Text Sources
Research Materials
