

Epitope-focused immunogens targeting the hepatitis C virus glycoproteins induce broadly neutralizing antibodies
- PMID: 39642219
- PMCID: PMC11623273
- DOI: 10.1126/sciadv.ado2600
Epitope-focused immunogens targeting the hepatitis C virus glycoproteins induce broadly neutralizing antibodies
Abstract
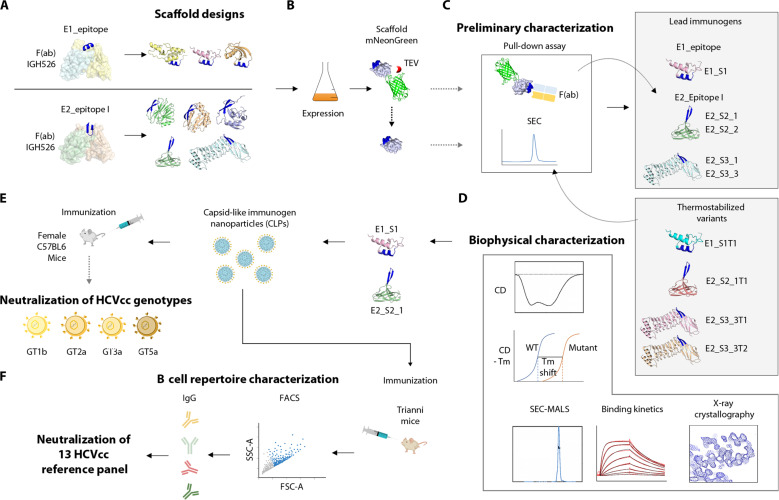
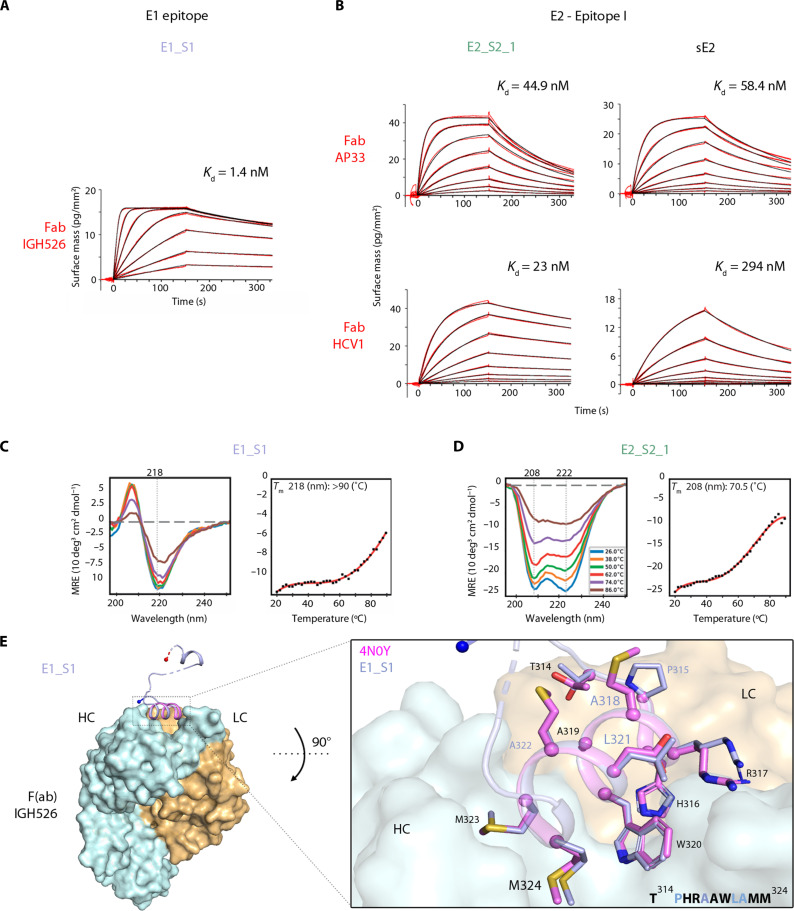
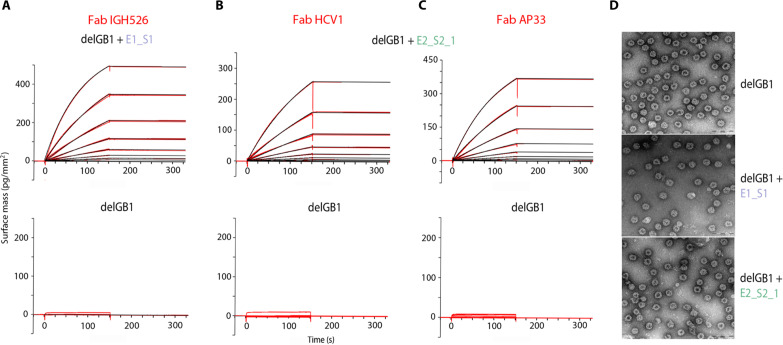
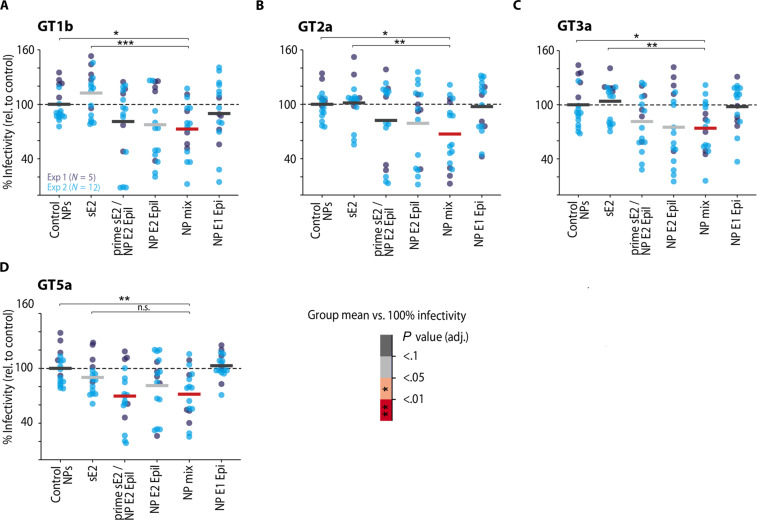
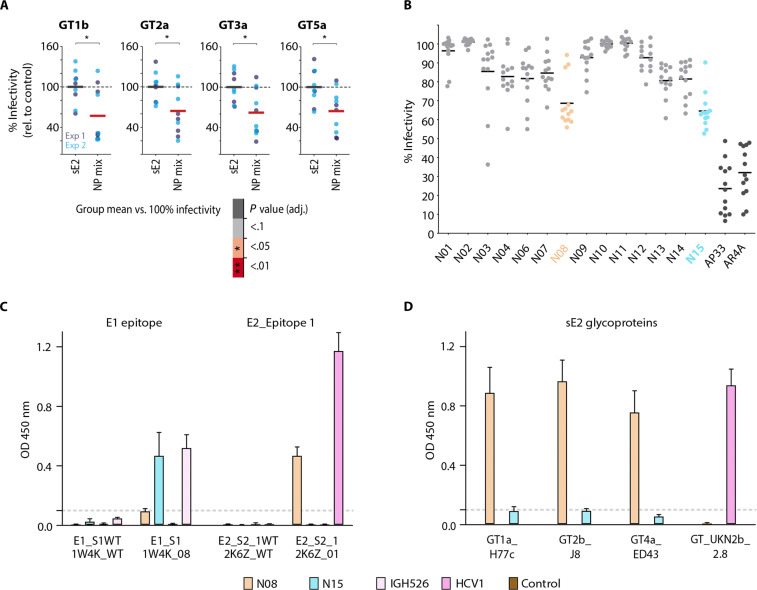
Hepatitis C virus (HCV) infection causes ~290,000 annual human deaths despite the highly effective antiviral treatment available. Several viral immune evasion mechanisms have hampered the development of an effective vaccine against HCV, among them the remarkable conformational flexibility within neutralization epitopes in the HCV antigens. Here, we report the design of epitope-focused immunogens displaying two distinct HCV cross-neutralization epitopes. We show that these immunogens induce a pronounced, broadly neutralizing antibody response in laboratory and transgenic human antibody mice. Monoclonal human antibodies isolated from immunized human antibody mice specifically recognized the grafted epitopes and neutralized four diverse HCV strains. Our results highlight a promising strategy for developing HCV immunogens and provide an encouraging paradigm for targeting structurally flexible epitopes to improve the induction of neutralizing antibodies.
Figures
References
-
- W. H. Organization, “Global Hepatitis Report 2021” (2021).
-
- Thimme R., T cell immunity to hepatitis C virus: Lessons for a prophylactic vaccine. J. Hepatol. 74, 220–229 (2021). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
