

Generating human neural diversity with a multiplexed morphogen screen in organoids
- PMID: 39642864
- PMCID: PMC12302009
- DOI: 10.1016/j.stem.2024.10.016
Generating human neural diversity with a multiplexed morphogen screen in organoids
Abstract
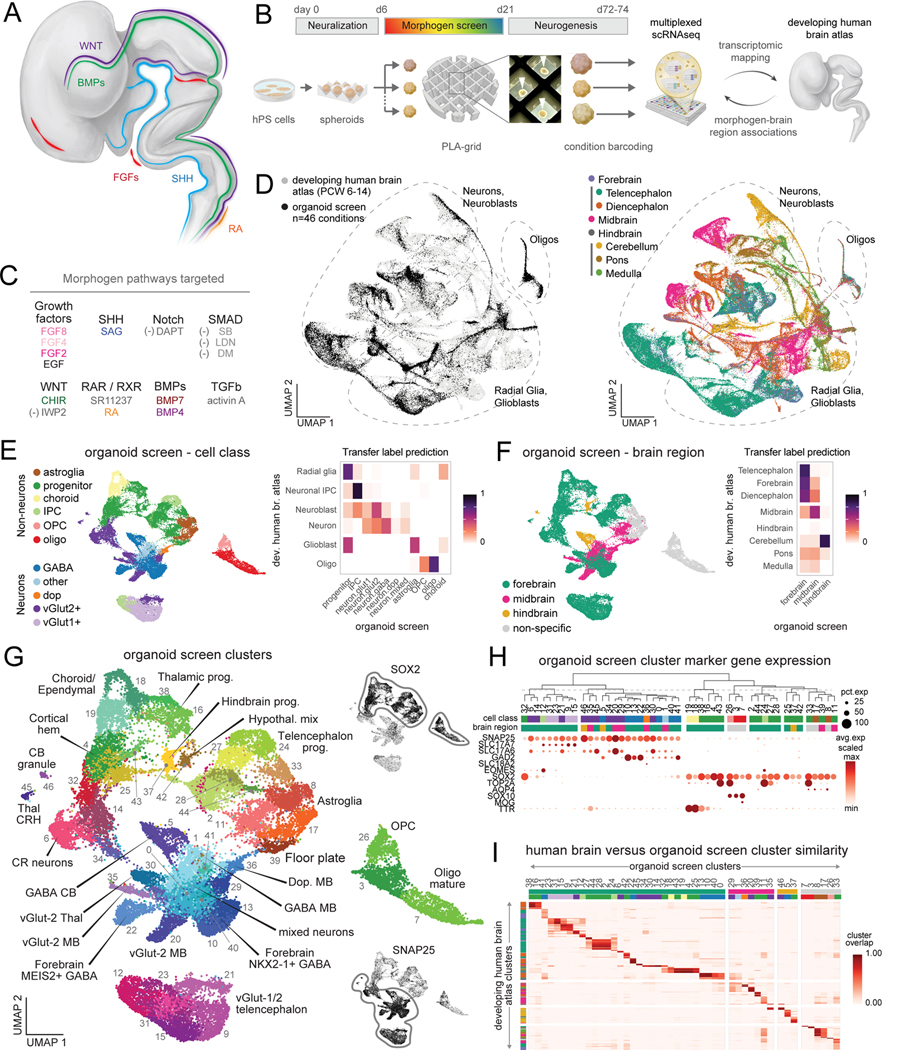
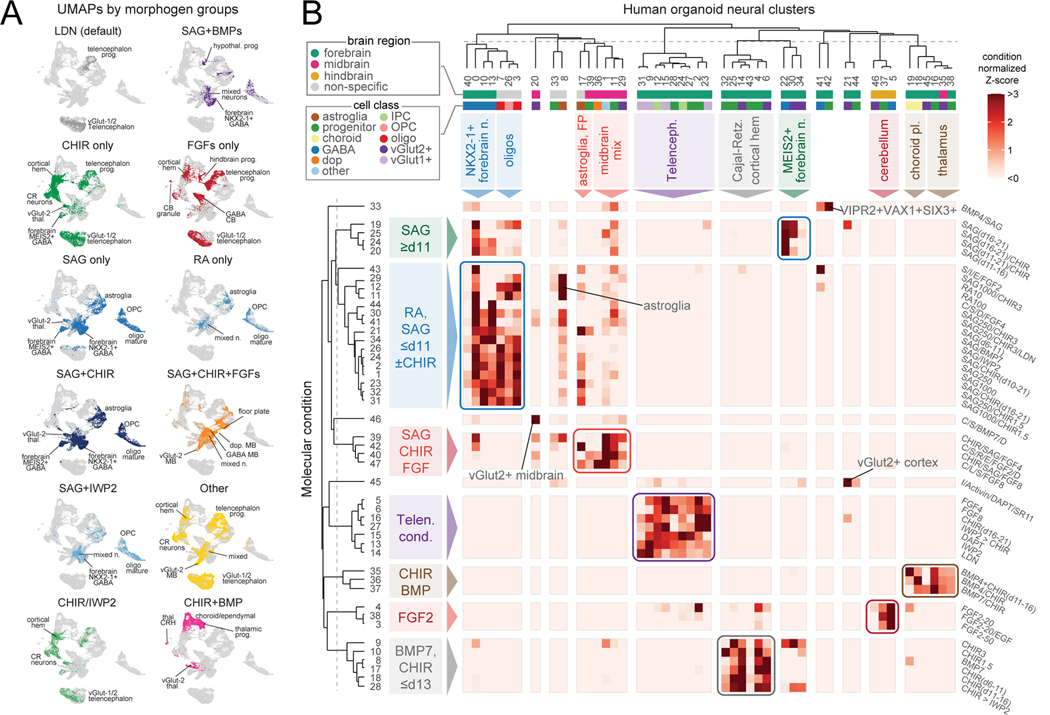
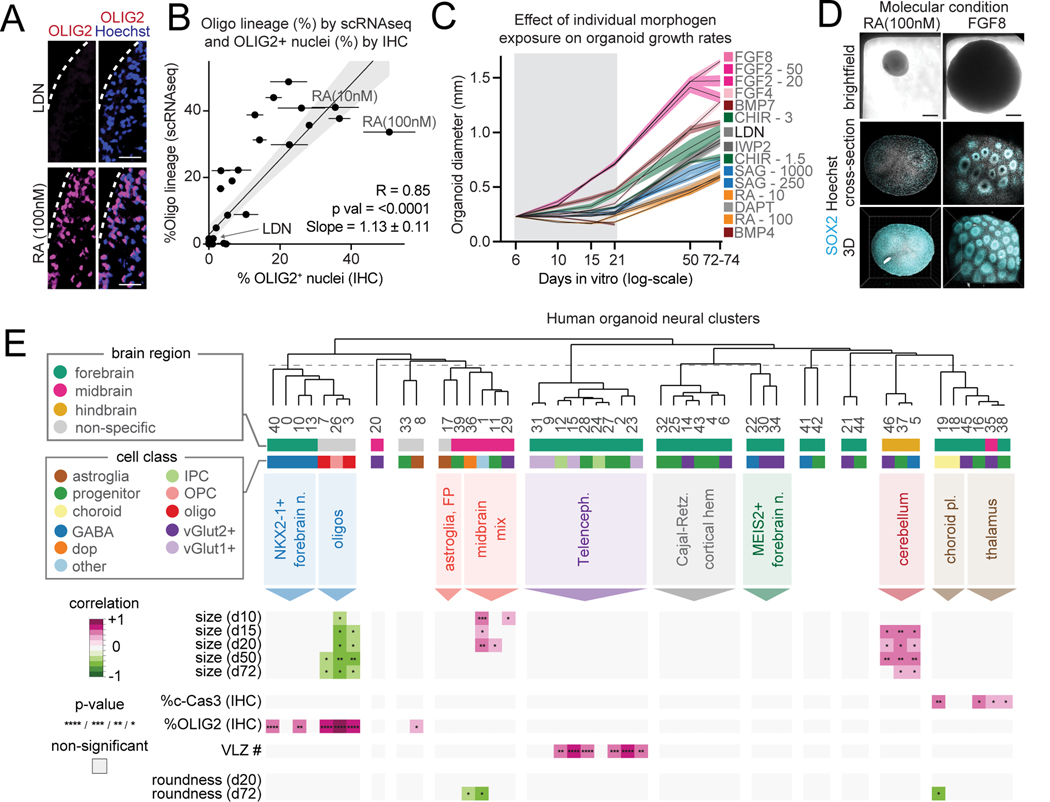
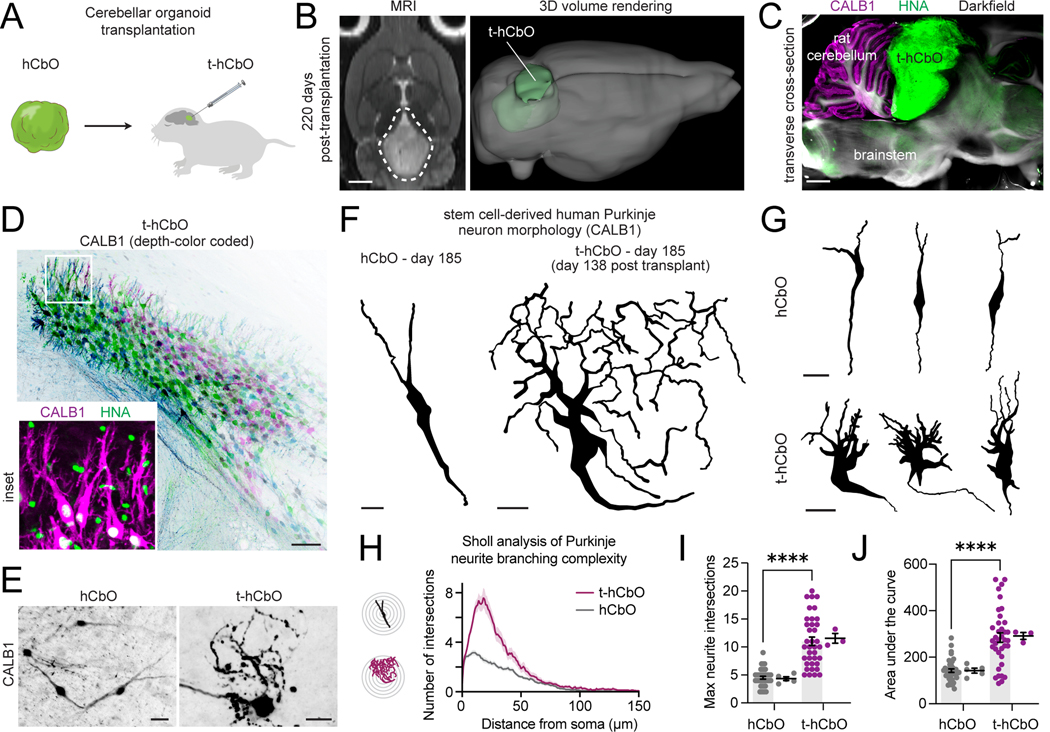
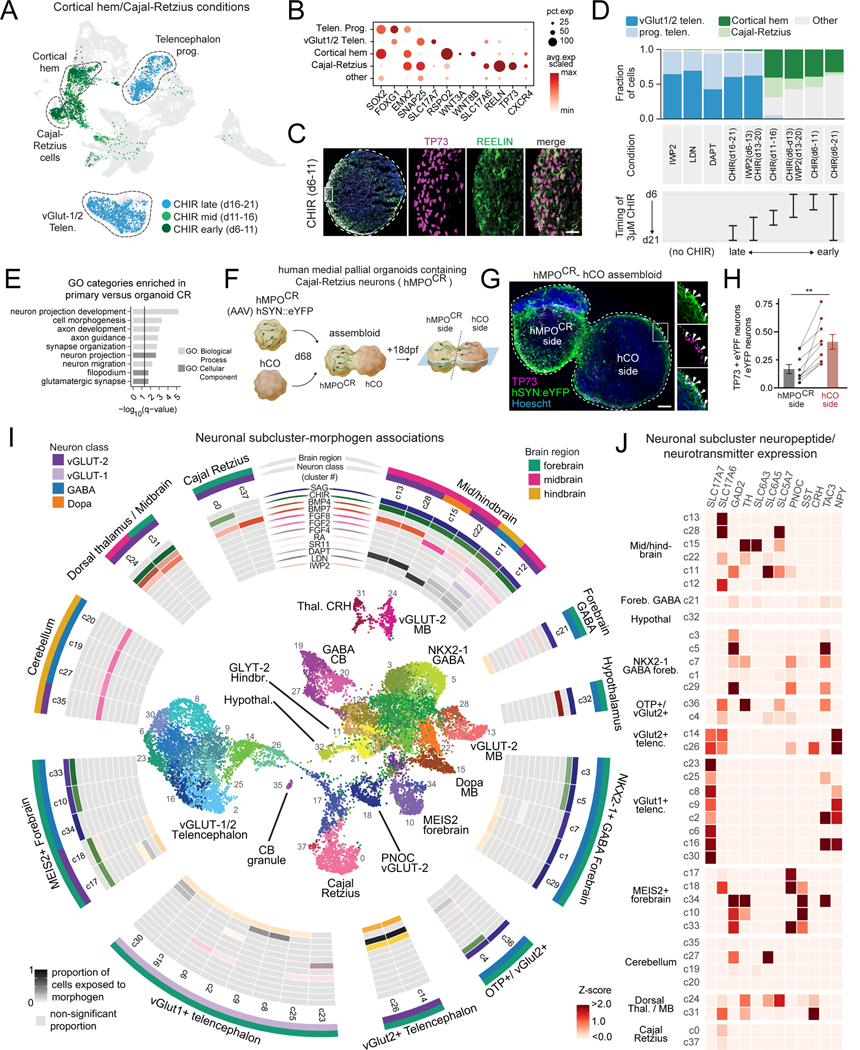
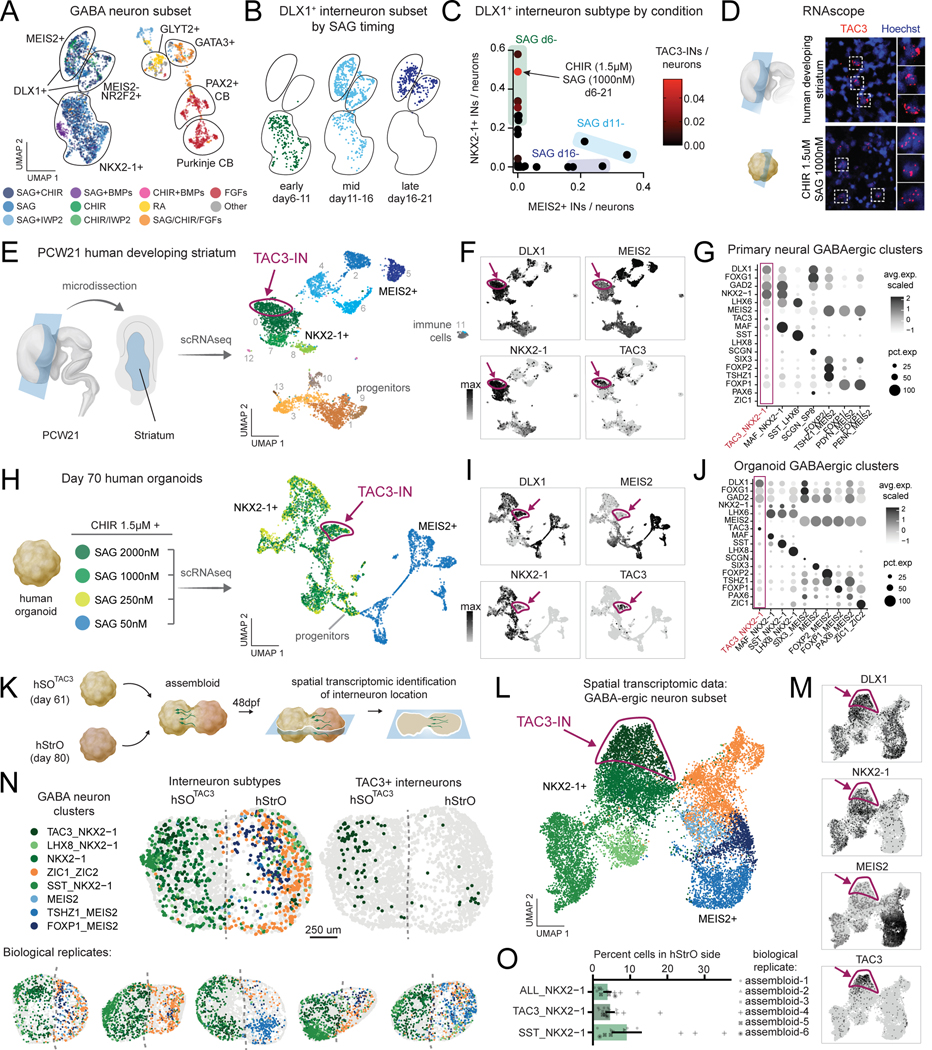
Morphogens choreograph the generation of remarkable cellular diversity in the developing nervous system. Differentiation of stem cells in vitro often relies upon the combinatorial modulation of these signaling pathways. However, the lack of a systematic approach to understand morphogen-directed differentiation has precluded the generation of many neural cell populations, and the general principles of regional specification and maturation remain incomplete. Here, we developed an arrayed screen of 14 morphogen modulators in human neural organoids cultured for over 70 days. Deconvolution of single-cell-multiplexed RNA sequencing data revealed design principles of brain region specification. We tuned neural subtype diversity to generate a tachykinin 3 (TAC3)-expressing striatal interneuron type within assembloids. To circumvent limitations of in vitro neuronal maturation, we used a neonatal rat transplantation strategy that enabled human Purkinje neurons to develop their hallmark complex dendritic branching. This comprehensive platform yields insights into the factors influencing stem cell-derived neural diversification and maturation.
Keywords: Purkinje neuron; morphogens; multiplexed screen; organoids; single-cell RNA-seq; transplantation.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.N. was an employee of Daiichi-Sankyo Co., Ltd, when performing the experiments for this study; however, the company did not have any input and interpretation on the design of experiments and the data. Stanford University holds patents that cover the generation of regionalized neural organoids.
Figures

Update of
-
Generating human neural diversity with a multiplexed morphogen screen in organoids.bioRxiv [Preprint]. 2023 Jun 1:2023.05.31.541819. doi: 10.1101/2023.05.31.541819. bioRxiv. 2023. Update in: Cell Stem Cell. 2024 Dec 5;31(12):1831-1846.e9. doi: 10.1016/j.stem.2024.10.016. PMID: 37398073 Free PMC article. Updated. Preprint.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
