

The disordered effector RipAO of Ralstonia solanacearum destabilizes microtubule networks in Nicotiana benthamiana cells
- PMID: 39645148
- PMCID: PMC11730531
- DOI: 10.1016/j.mocell.2024.100167
The disordered effector RipAO of Ralstonia solanacearum destabilizes microtubule networks in Nicotiana benthamiana cells
Abstract
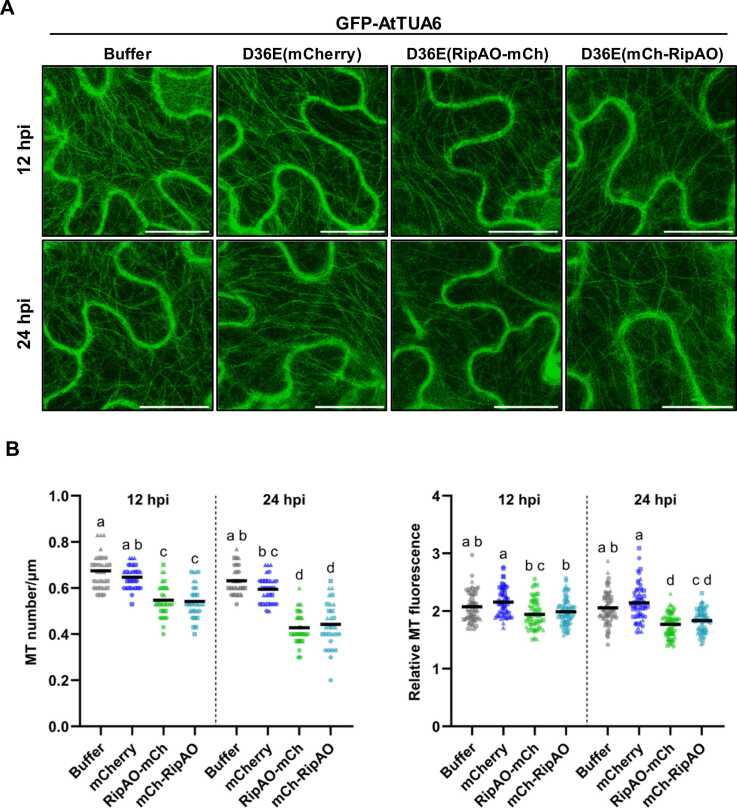
Ralstonia solanacearum causes bacterial wilt, a devastating disease in solanaceous crops. The pathogenicity of R. solanacearum depends on its type III secretion system, which delivers a suite of type III effectors into plant cells. The disordered core effector RipAO is conserved across R. solanacearum species and affects plant immune responses when transiently expressed in Nicotiana benthamiana. Specifically, RipAO impairs pathogen-associated molecular pattern-triggered reactive oxygen species production, an essential plant defense mechanism. RipAO fused to yellow fluorescent protein initially localizes to filamentous structures, resembling the cytoskeleton, before forming large punctate aggregates around the nucleus. Consistent with these findings, tubulin alpha 6 (TUA6) and tubulin beta-1, building blocks of microtubules, were identified as putative targets of RipAO in immunoprecipitation and mass spectrometry analyses. In the presence of RipAO, TUA6-labeled microtubules fragmented into puncta, mimicking the effects of oryzalin, a microtubule polymerization inhibitor. These effects were corroborated in a N. benthamiana transgenic line constitutively expressing green fluorescent protein-labeled TUA6, where RipAO reduced microtubule density and stability at an accumulation level that did not induce aggregation. Moreover, oryzalin treatment further enhanced RipAO's impairment of reactive oxygen species production, suggesting that RipAO disrupts microtubule networks via its association with tubulins, leading to immune suppression. Further research into RipAO's interaction with the microtubule network will enhance our understanding of bacterial strategies to subvert plant immunity.
Keywords: Cytoskeleton; Heterologous expression; Reactive oxygen species; Type III secreted effector; Virulence.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interests The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures






References
-
- Brandizzi F., Wasteneys G.O. Cytoskeleton-dependent endomembrane organization in plant cells: an emerging role for microtubules. Plant J. 2013;75:339–349. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
