

TMEM135 deficiency improves hepatic steatosis by suppressing CD36 in a SIRT1-dependent manner
- PMID: 39647810
- PMCID: PMC11728970
- DOI: 10.1016/j.molmet.2024.102080
TMEM135 deficiency improves hepatic steatosis by suppressing CD36 in a SIRT1-dependent manner
Abstract
Objectives: Dysregulation of lipid homeostasis pathway causes many liver diseases, including hepatic steatosis. One of the primary factors contributing to lipid accumulation is fatty acid uptake by the liver. Transmembrane protein 135 (TMEM135), which exists in mitochondria and peroxisomes, participates in intracellular lipid metabolism. This study aims to investigate the role of TMEM135 on regulating cellular lipid import in the liver.
Methods: We used in vivo, ex vivo, and in vitro models of steatosis. TMEM135 knockout (TMEM135KO) and wild type (WT) mice were fed a high-fat diet (HFD) to induce hepatic steatosis. Primary mouse hepatocytes and AML12 cells were treated with free fatty acid (FFA). Additionally, TMEM135-deficient stable cells and overexpressed cells were established using AML12 cells.
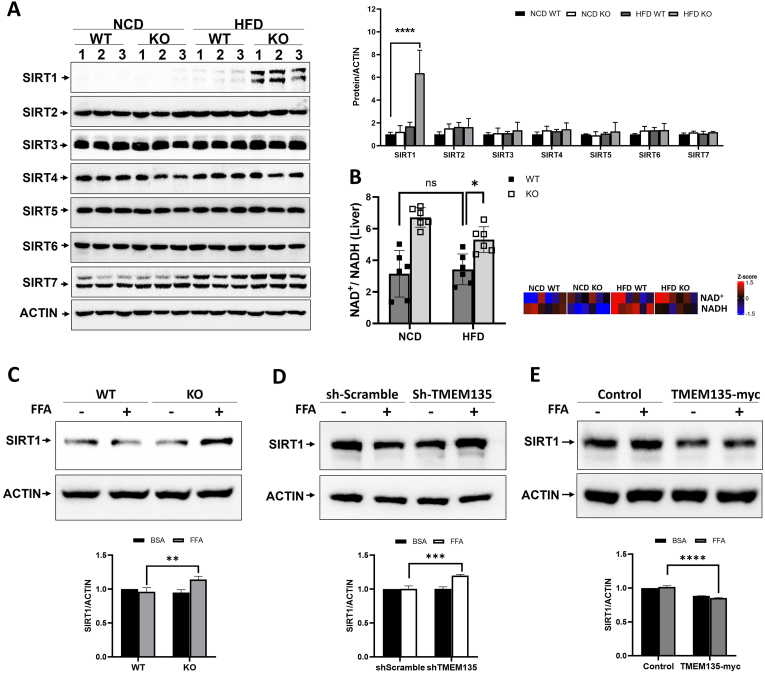
Results: TMEM135 deficiency mitigated lipid accumulation in the liver of HFD-fed TMEM135KO mice. TMEM135-depleted primary hepatocytes and AML12 cells exhibited less lipid accumulation when treated with FFA compared to control cells, as shown as lipid droplets. Consistently, the effect of TMEM135 depletion on lipid accumulation was completely reversed under TMEM135 overexpression conditions. CD36 expression was markedly induced by HFD or FFA, which was reduced by TMEM135 depletion. Among the SIRT family proteins, only SIRT1 expression definitely increased in the liver of HFD-fed TMEM135KO mice along with a significant increase in NAD+/NADH ratio. However, inhibition of SIRT1 in TMEM135-depleted cells using siSIRT1 or the SIRT1 inhibitor EX-527 resulted in an increase of CD36 expression and consequent TG levels.
Conclusions: TMEM135 depletion attenuates CD36 expression in a SIRT1-dependent manner, thereby reducing cellular lipid uptake and hepatic steatosis.
Keywords: CD36; Lipid accumulation; SIRT1; TMEM135.
Copyright © 2024. Published by Elsevier GmbH.
Conflict of interest statement
Declaration of competing interest The authors who have taken part in this study declared that they do not have anything to disclose regarding funding or conflict of interest with respect to this manuscript.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
