

Hsp90 and cochaperones have two genetically distinct roles in regulating eEF2 function
- PMID: 39652595
- PMCID: PMC11651573
- DOI: 10.1371/journal.pgen.1011508
Hsp90 and cochaperones have two genetically distinct roles in regulating eEF2 function
Abstract
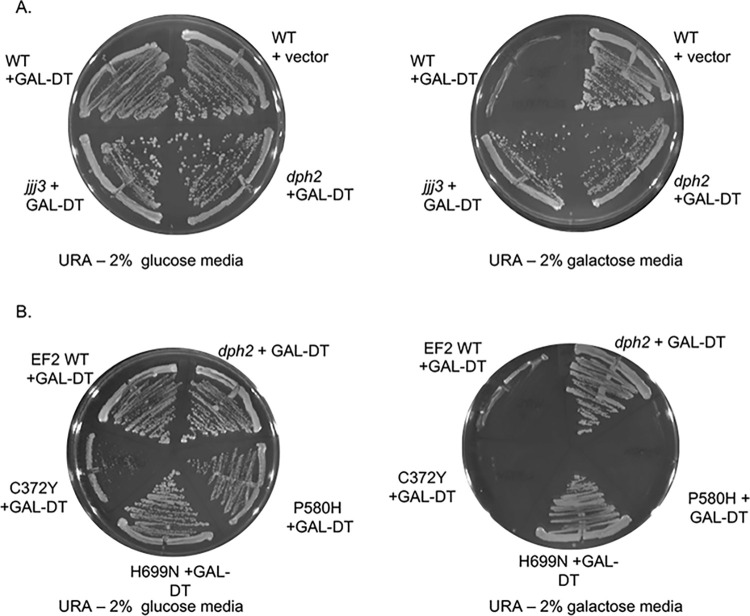
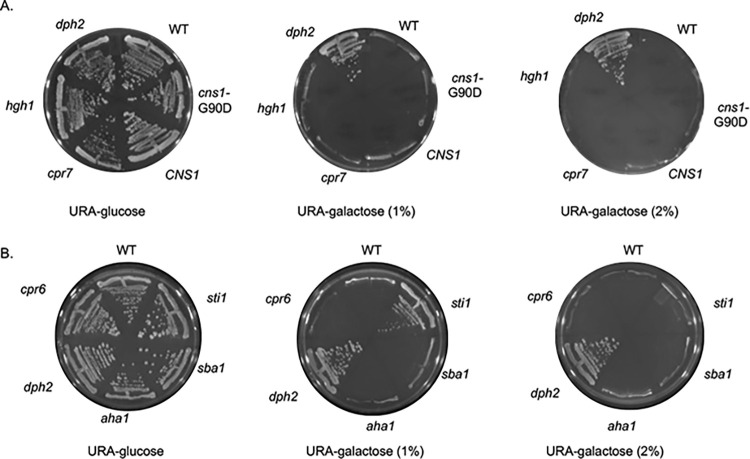
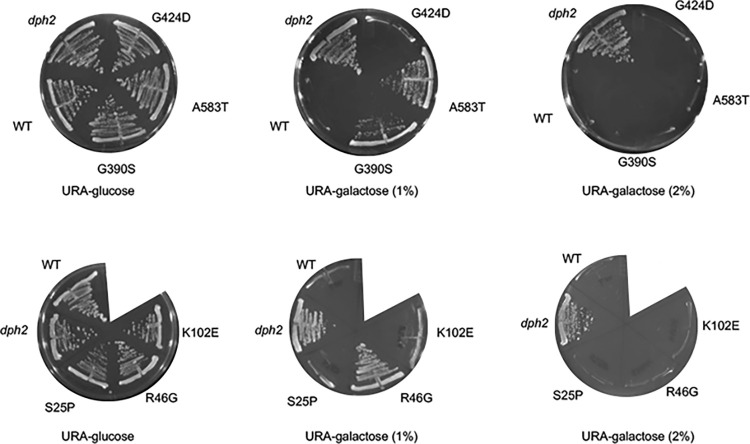
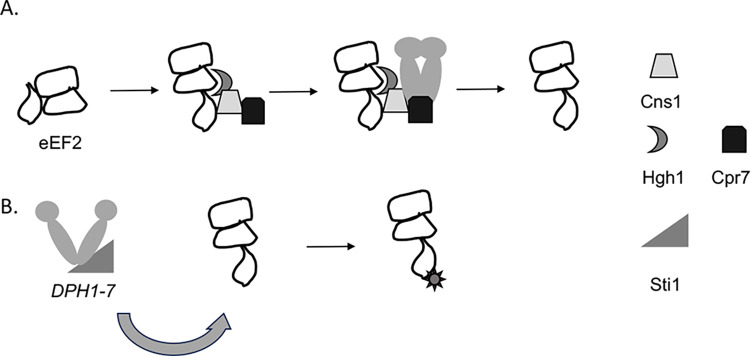
Protein homeostasis relies on the accurate translation and folding of newly synthesized proteins. Eukaryotic elongation factor 2 (eEF2) promotes GTP-dependent translocation of the ribosome during translation. eEF2 folding was recently shown to be dependent on Hsp90 as well as the cochaperones Hgh1, Cns1, and Cpr7. We examined the requirement for Hsp90 and cochaperones more closely and found that Hsp90 and cochaperones have two distinct roles in regulating eEF2 function. Yeast expressing one group of Hsp90 mutations or one group of cochaperone mutations had reduced steady-state levels of eEF2. The growth of Hsp90 mutants that affected eEF2 accumulation was also negatively affected by deletion of the gene encoding Hgh1. Further, mutations in yeast eEF2 that mimic disease-associated mutations in human eEF2 were negatively impacted by loss of Hgh1 and growth of one mutant was partially rescued by overexpression of Hgh1. In contrast, yeast expressing different groups of Hsp90 mutations or a different cochaperone mutation had altered sensitivity to diphtheria toxin, which is dictated by a unique posttranslational modification on eEF2. Our results provide further evidence that Hsp90 contributes to proteostasis not just by assisting protein folding, but also by enabling accurate translation of newly synthesized proteins. In addition, these results provide further evidence that yeast Hsp90 mutants have distinct in vivo effects that correlate with defects in subsets of cochaperones.
Copyright: © 2024 Fulton et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
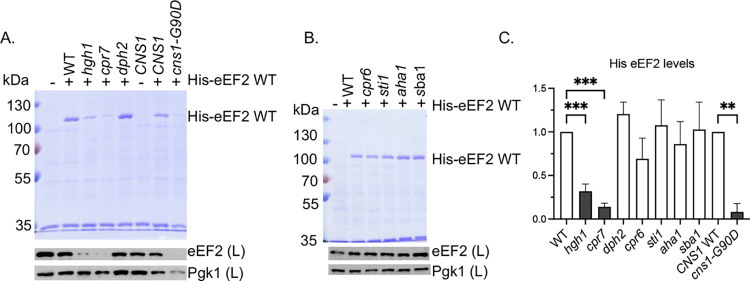
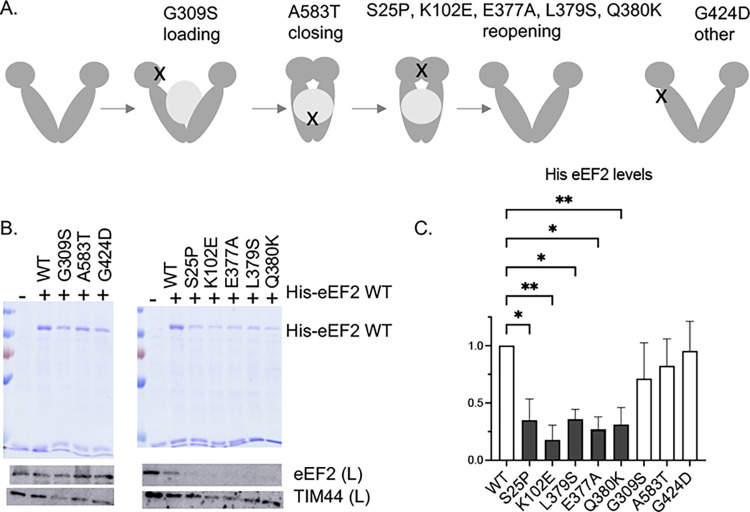
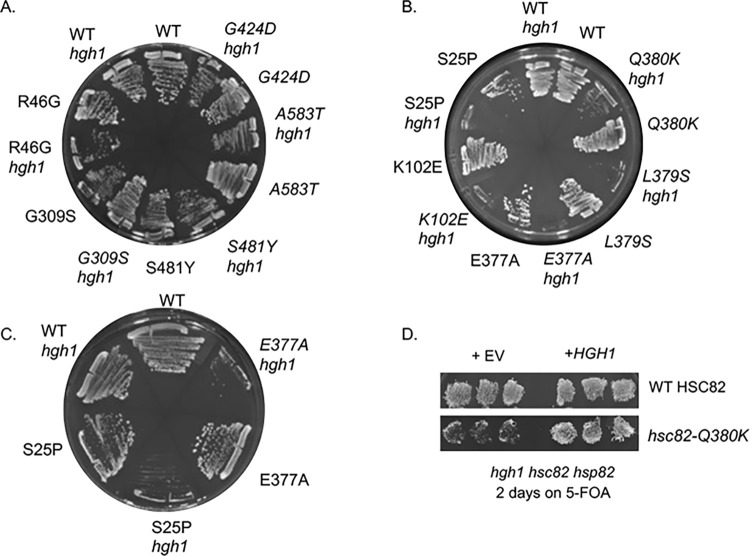
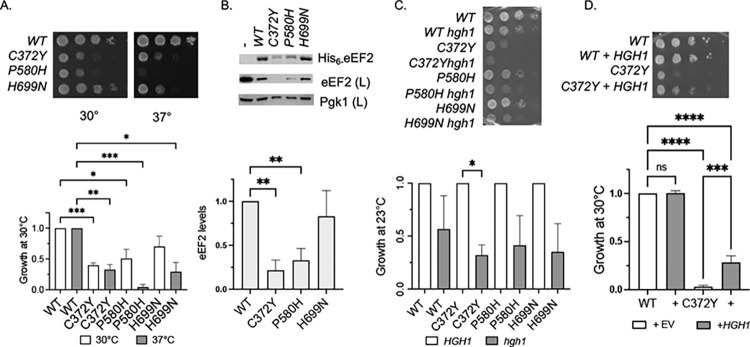
Similar articles
-
The Co-chaperone Cns1 and the Recruiter Protein Hgh1 Link Hsp90 to Translation Elongation via Chaperoning Elongation Factor 2.Mol Cell. 2019 Apr 4;74(1):73-87.e8. doi: 10.1016/j.molcel.2019.02.011. Epub 2019 Mar 12. Mol Cell. 2019. PMID: 30876805
-
Diphthamide synthesis is linked to the eEF2-client chaperone machinery.FEBS Lett. 2025 May;599(9):1260-1268. doi: 10.1002/1873-3468.15095. Epub 2025 Jan 17. FEBS Lett. 2025. PMID: 39825589 Free PMC article.
-
The Hsp90 cochaperones Cpr6, Cpr7, and Cns1 interact with the intact ribosome.Eukaryot Cell. 2015 Jan;14(1):55-63. doi: 10.1128/EC.00170-14. Epub 2014 Nov 7. Eukaryot Cell. 2015. PMID: 25380751 Free PMC article.
-
The Hsp70-Hsp90 Chaperone Cascade in Protein Folding.Trends Cell Biol. 2019 Feb;29(2):164-177. doi: 10.1016/j.tcb.2018.10.004. Epub 2018 Nov 28. Trends Cell Biol. 2019. PMID: 30502916 Review.
-
Evolution and function of diverse Hsp90 homologs and cochaperone proteins.Biochim Biophys Acta. 2012 Mar;1823(3):607-13. doi: 10.1016/j.bbamcr.2011.09.020. Epub 2011 Oct 8. Biochim Biophys Acta. 2012. PMID: 22008467 Review.
Cited by
-
Hsp90: Bringing it all together.Cell Stress Chaperones. 2025 Feb;30(1):69-79. doi: 10.1016/j.cstres.2025.01.002. Epub 2025 Jan 29. Cell Stress Chaperones. 2025. PMID: 39889818 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
