

Galectin-3 plays a key role in controlling infection by Toxoplasma gondii in human trophoblast cells and human villous explants
- PMID: 39654979
- PMCID: PMC11625798
- DOI: 10.3389/fcimb.2024.1459810
Galectin-3 plays a key role in controlling infection by Toxoplasma gondii in human trophoblast cells and human villous explants
Abstract
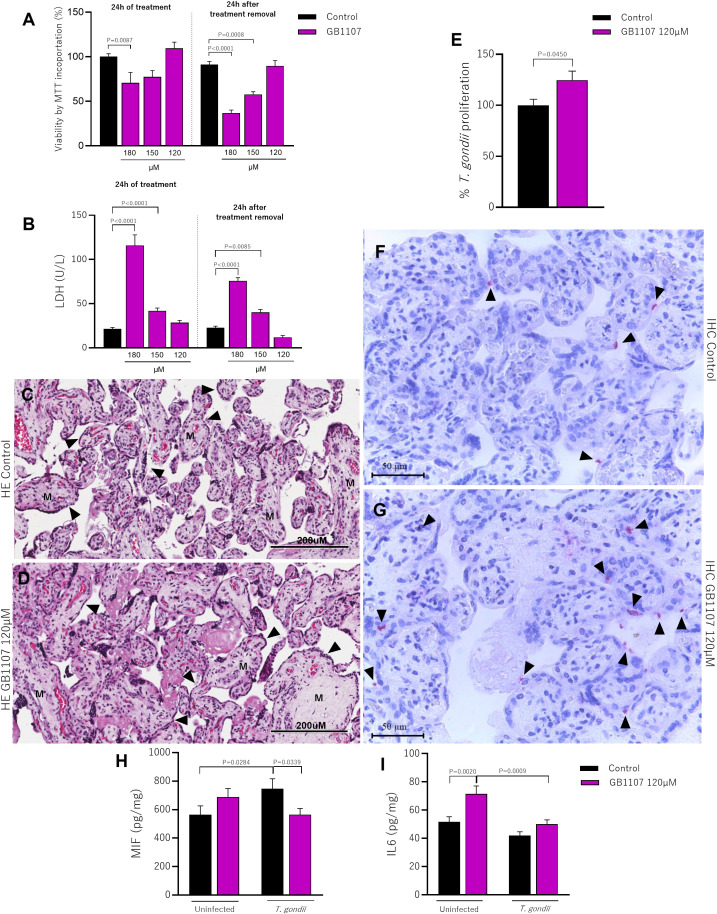
Galectin-3 (Gal-3) is a β-galactoside-binding lectin expressed in cells of the placental microenvironment. This lectin is involved in various biological processes, such as modulation of the immune system and control of parasitic illness. Toxoplasma gondii infection can lead to congenital transmission and cause miscarriages, prematurity and fetal anomalies. However, little is known about the role of Gal-3 in T. gondii infection in the placental microenvironment. This study aimed to unravel the underlying mechanisms of Gal-3 during T. gondii infection. For this purpose, we promoted the knockdown of Gal-3 expression by using RNA interference (RNAi) in BeWo cells or by using a synthetic inhibitor (GB1107) in human villous explants. We showed that the decreased Gal-3 expression in BeWo cells and human villous explants increases the invasion and proliferation of T. gondii probably by downregulating MIF and IL6 levels, highlighting thus the role of this lectin in modulating the immune response. Collectively, our study reveals Gal-3 as a promising target protein during congenital toxoplasmosis.
Keywords: Gal-3; congenital toxoplasmosis; immune response; maternal-fetal interface; placental.
Copyright © 2024 Luz, Ribeiro, Teixeira, de Souza, Paschoalino, Sousa, Rosini, dos Santos, de Oliveira, de Lima Júnior, Damasceno, Almeida, Barbosa, Alves, da Silva, Barbosa and Ferro.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



References
-
- Alves C. M., Silva D. A., Azzolini A. E., Marzocchi-MaChado C. M., Carvalho J. V., Pajuaba A. C., et al. . (2010). Galectin-3 plays a modulatory role in the life span and activation of murine neutrophils during early Toxoplasma gondii infection. Immunobiology 215, 475–485. doi: 10.1016/j.imbio.2009.08.001 - DOI - PubMed
-
- Alves C. M., Silva D. A., Azzolini A. E., Marzocchi-MaChado C. M., Lucisano-Valim Y. M., Roque-Barreira M. C., et al. . (2013). Galectin-3 is essential for reactive oxygen species production by peritoneal neutrophils from mice infected with a virulent strain of Toxoplasma gondii. Parasitology 140, 210–219. doi: 10.1017/s0031182012001473 - DOI - PubMed
-
- Barbosa B. F., Lopes-Maria J. B., Gomes A. O., Angeloni M. B., Castro A. S., Franco P. S., et al. . (2015). IL10, TGF beta1, and IFN gamma modulate intracellular signaling pathways and cytokine production to control Toxoplasma gondii infection in BeWo trophoblast cells. Biol. Reprod. 92, 82. doi: 10.1095/biolreprod.114.124115 - DOI - PubMed
-
- Bernardes E. S., Silva N. M., Ruas L. P., Mineo J. R., Loyola A. M., Hsu D. K., et al. . (2006). Toxoplasma gondii infection reveals a novel regulatory role for galectin-3 in the interface of innate and adaptive immunity. Am. J. Pathol. 168, 1910–1920. doi: 10.2353/ajpath.2006.050636 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous