

Modeling the within-host dynamics of Plasmodium vivax hypnozoite activation: An analysis of the SPf66 vaccine trial
- PMID: 39656209
- PMCID: PMC11665876
- DOI: 10.1073/pnas.2401024121
Modeling the within-host dynamics of Plasmodium vivax hypnozoite activation: An analysis of the SPf66 vaccine trial
Abstract
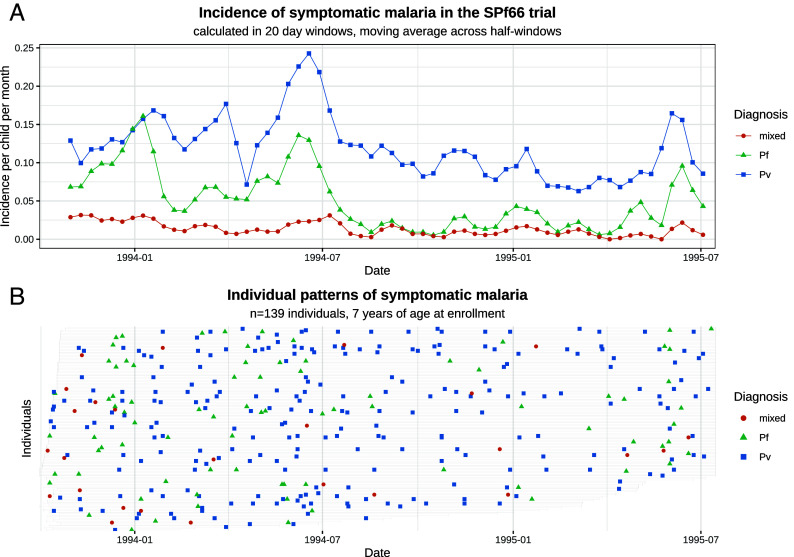
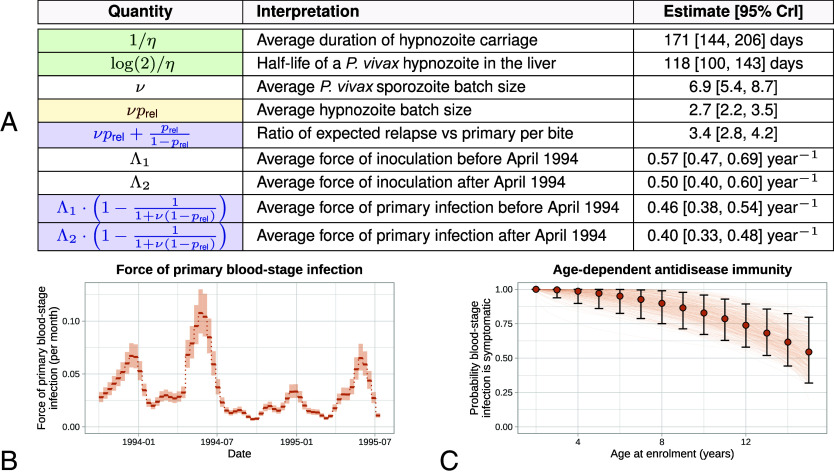
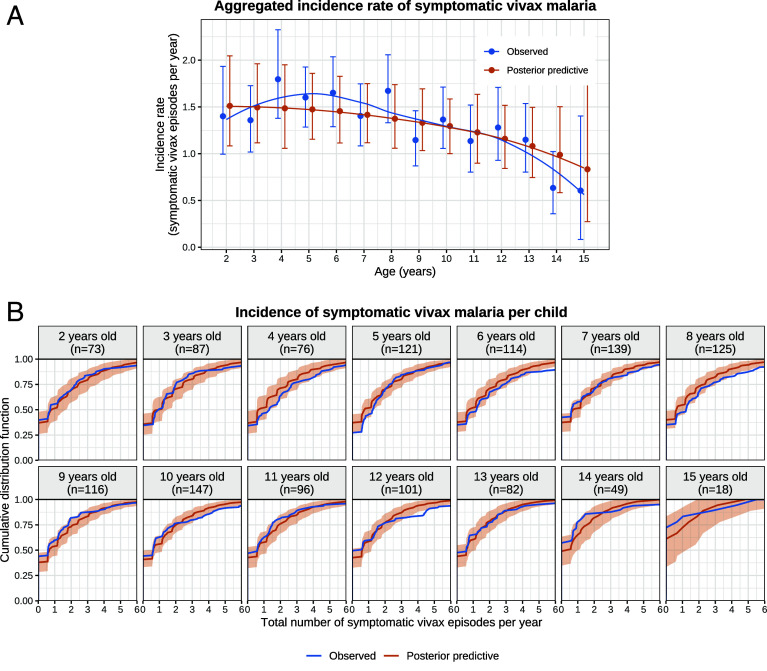
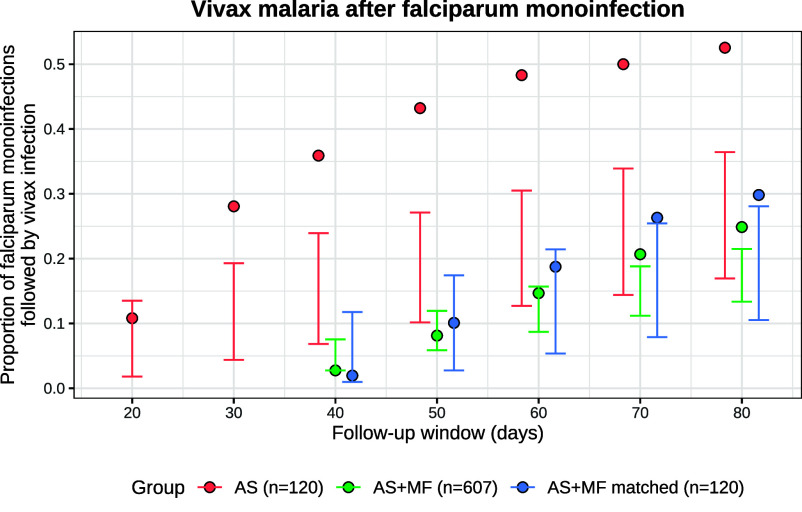
Plasmodium vivax parasites can lie dormant in the liver as hypnozoites, activating weeks to months after sporozoite inoculation to cause relapsing malarial illness. It is not known what biological processes govern hypnozoite activation. We use longitudinal data from the most detailed cohort study ever conducted in an area where both Plasmodium falciparum and P. vivax were endemic to fit a simple within-host mathematical model of P. vivax hypnozoite activation. 1,344 children living on the Thailand-Myanmar border were followed daily for 21 mo. There were 2,504 vivax and 1,164 falciparum malaria symptomatic episodes recorded over 1988 person-years. The model assumes that hypnozoites activate independently at a constant rate ("exponential clock model"). When this model was embedded in a stochastic framework for repeated infectious mosquito bites, with seasonality inferred from the incidence of clinical falciparum malaria episodes, it explained the observed temporal patterns of multiple (up to 13) recurrent vivax malaria episodes. Under this model, we estimate the mean dormancy period for a single hypnozoite to be 6 mo (i.e., a half-life of 4 mo). We use the calibrated within-host model to characterize population-level overdispersion in the risk of relapse, and assess the potential utility of a serological test for radical cure in low transmission settings. We show that mefloquine treatment of falciparum malaria eliminates early vivax relapses; and that there are substantially more P. vivax recurrences than expected under the model following artesunate monotherapy treatment for falciparum malaria. These results suggest that hypnozoites can be activated by symptomatic malarial illness.
Keywords: exponential clock; hypnozoite; mathematical model; vivax malaria; within-host.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
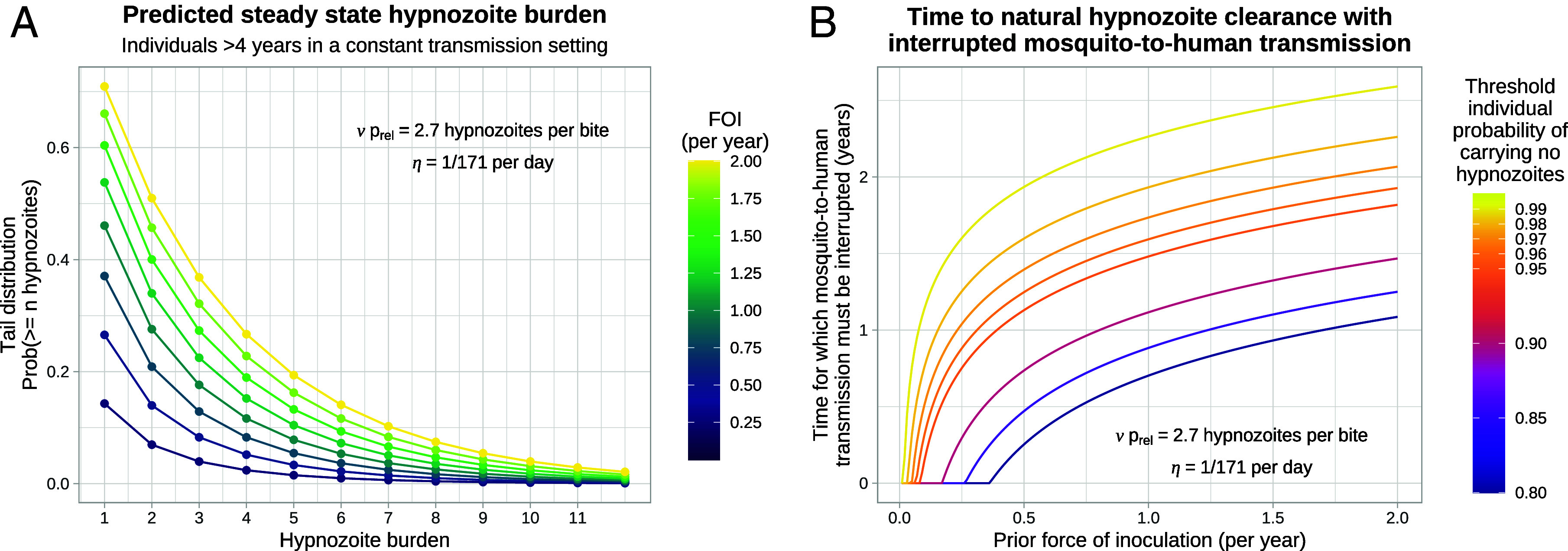
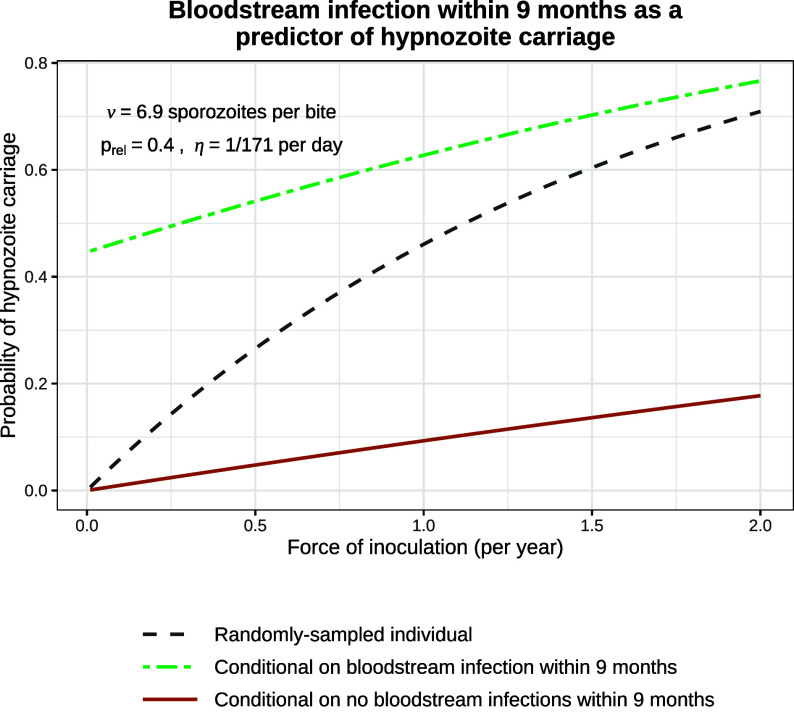
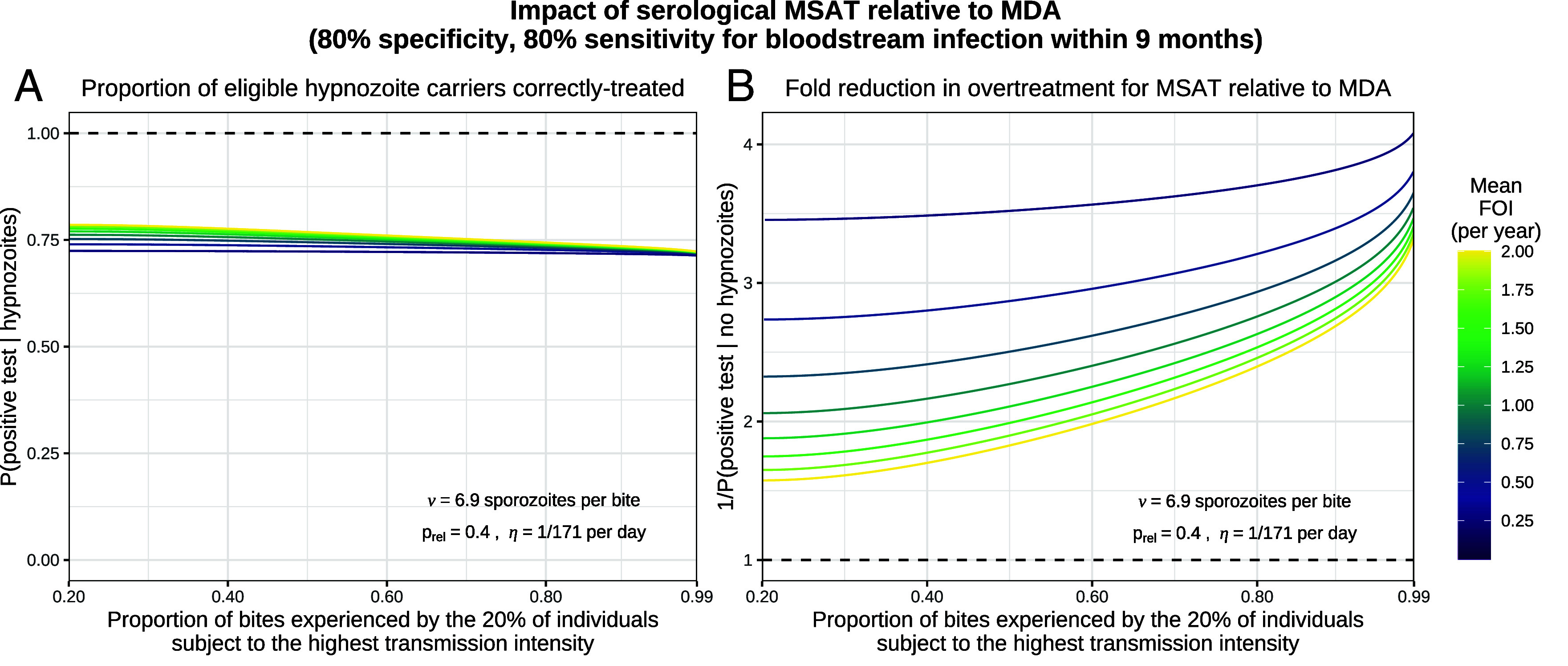
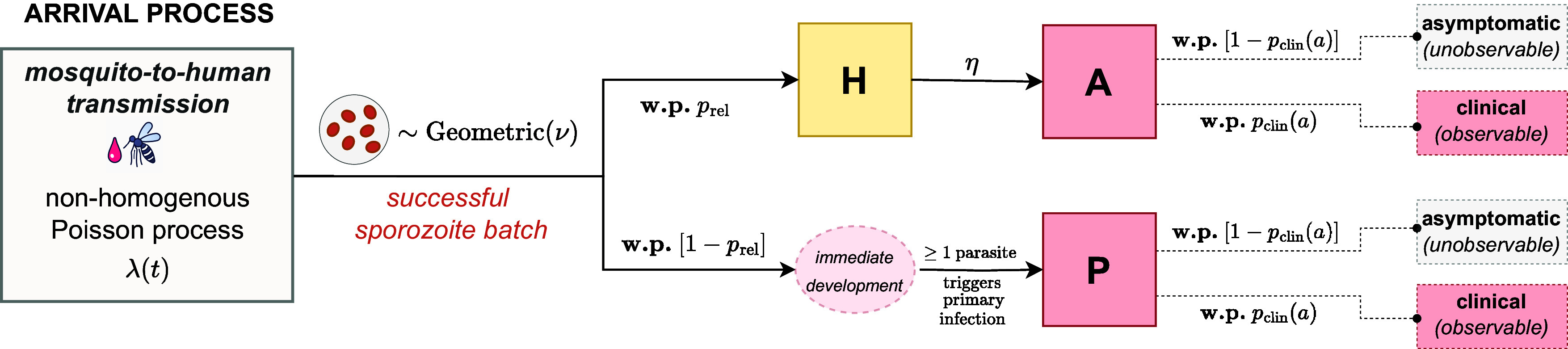
References
-
- World Health Organization, WHO Guidelines for Malaria (World Health Organization, 2023).
-
- Schäfer C., Zanghi G., Vaughan A. M., Kappe S. H., Plasmodium vivax latent liver stage infection and relapse: Biological insights and new experimental tools. Annu. Rev. Microbiol. 75, 87–106 (2021). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
