

Rooting for survival: how plants tackle a challenging environment through a diversity of root forms and functions
- PMID: 39657006
- PMCID: PMC11663570
- DOI: 10.1093/plphys/kiae586
Rooting for survival: how plants tackle a challenging environment through a diversity of root forms and functions
Abstract
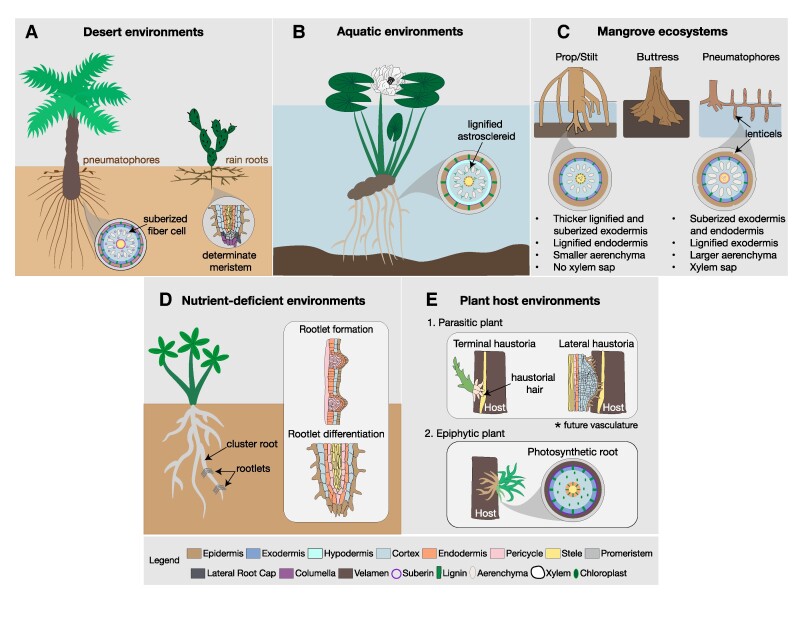
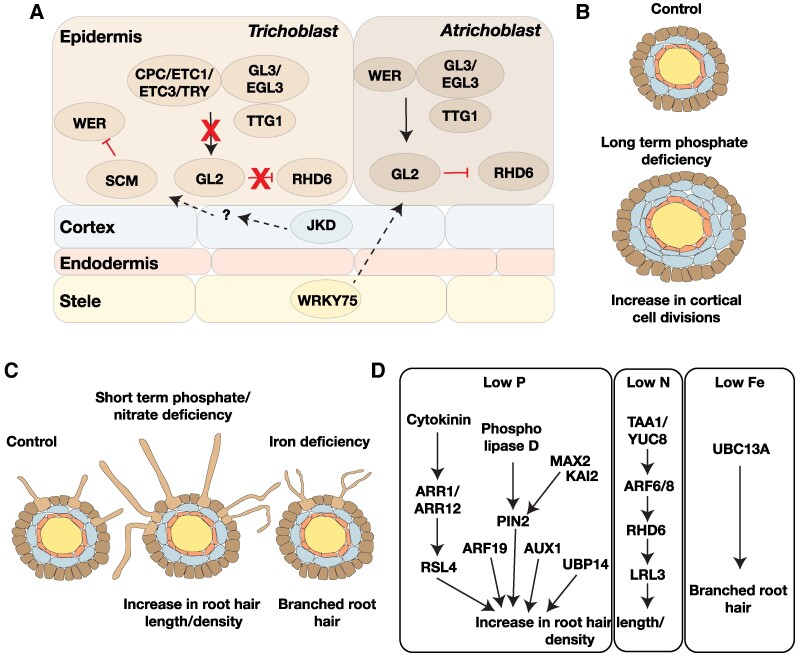
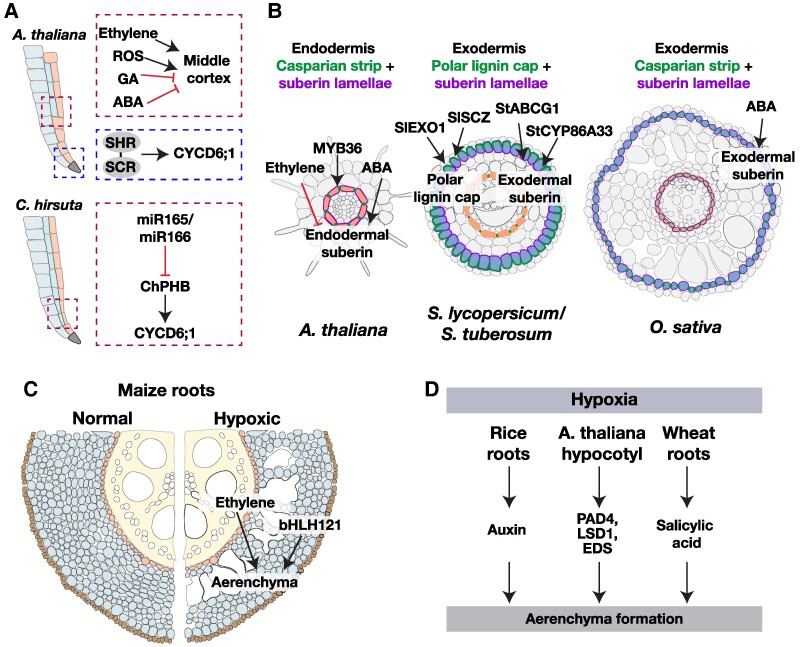
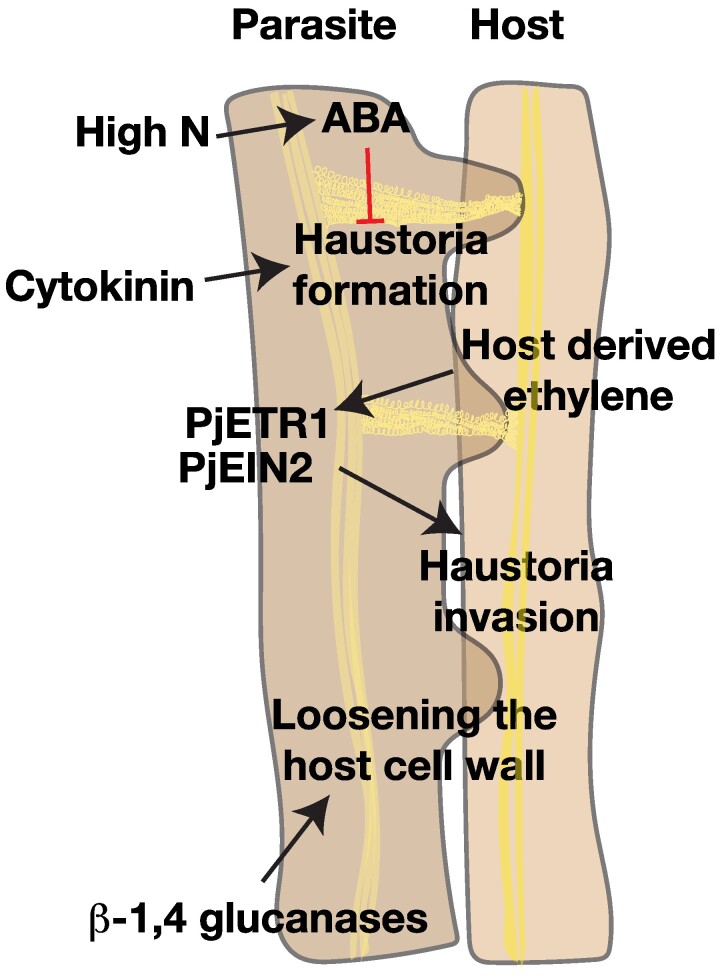
The current climate crisis has global impacts and will affect the physiology of plants across every continent. Ensuring resilience of our agricultural and natural ecosystems to the environmental stresses imposed by climate change will require molecular insight into the adaptations employed by a diverse array of plants. However, most current studies continue to focus on a limited set of model species or crops. Root systems are particularly understudied even though their functions in water and nutrient uptake are likely pivotal for plant stress resilience and sustainable agriculture. In this review, we highlight anatomical adaptations in roots that enable plant survival in different ecological niches. We then present the current state of knowledge for the molecular underpinnings of these adaptations. Finally, we identify areas where future research using a biodiversity approach can fill knowledge gaps necessary for the development of climate-resilient crops of the future.
© The Author(s) 2024. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures
References
-
- Abd Elhalim ME, Abo-Alatta O, Habib SA, Abd Elbar OH. The anatomical features of the desert halophytes Zygophyllum album L.F. and Nitraria retusa (Forssk.) Asch. Ann Agric Sci. 2016:61(1):97–104. 10.1016/j.aoas.2015.12.001 - DOI
-
- Ameloot E, Verheyen K, Hermy M. Meta-analysis of standing crop reduction by Rhinanthus spp. and its effect on vegetation structure. Folia Geobot. 2005:40(2–3):289–310. 10.1007/BF02803241 - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
