

Enhanced fatty acid oxidation in osteoprogenitor cells provides protection from high-fat diet induced bone dysfunction
- PMID: 39657629
- PMCID: PMC11789392
- DOI: 10.1093/jbmr/zjae195
Enhanced fatty acid oxidation in osteoprogenitor cells provides protection from high-fat diet induced bone dysfunction
Abstract
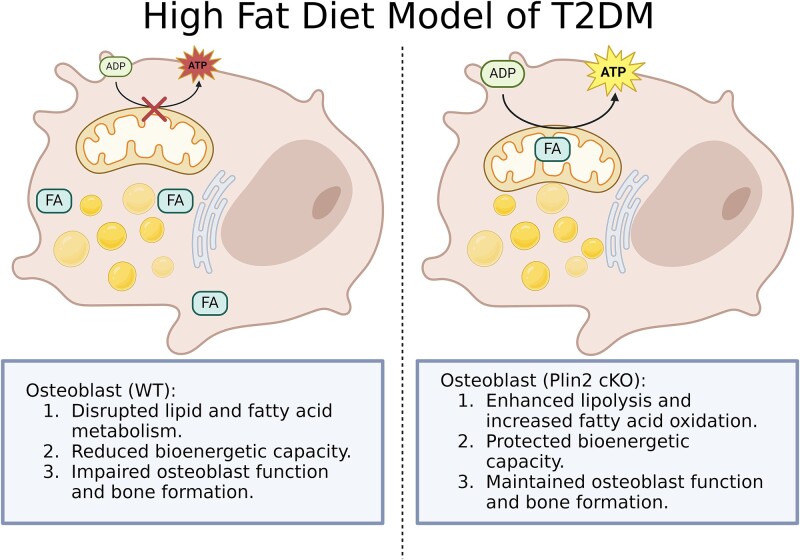
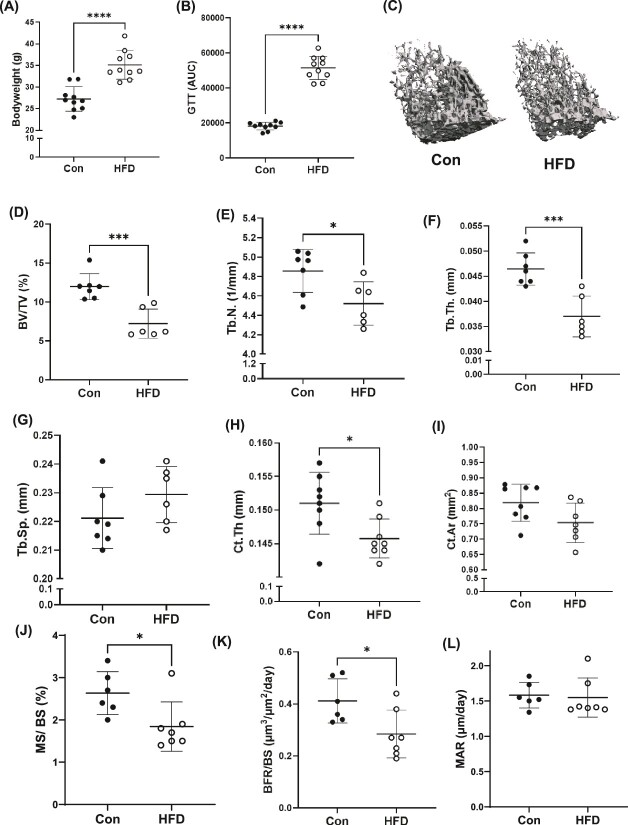
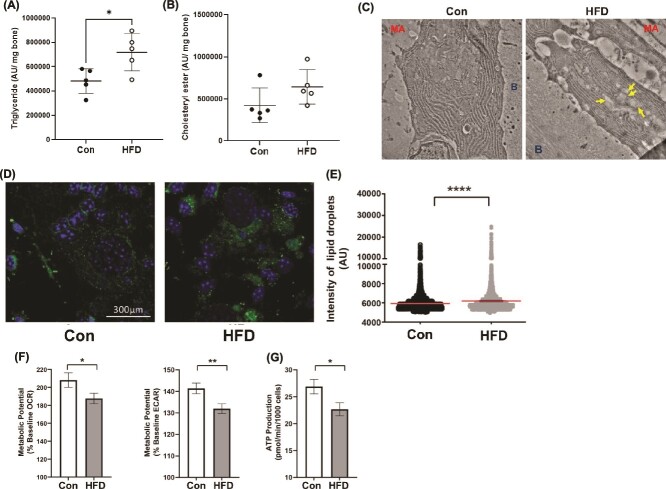
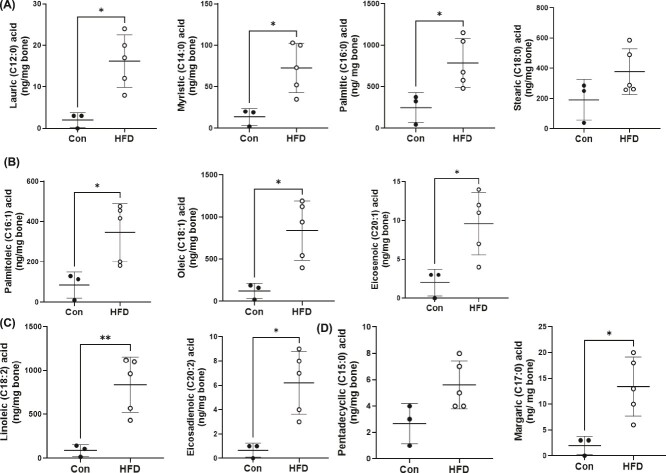
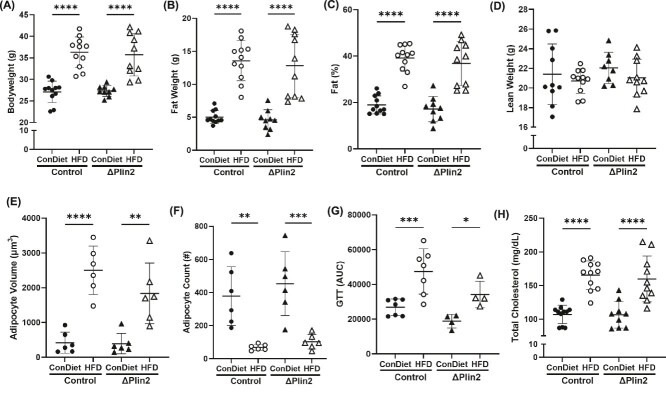
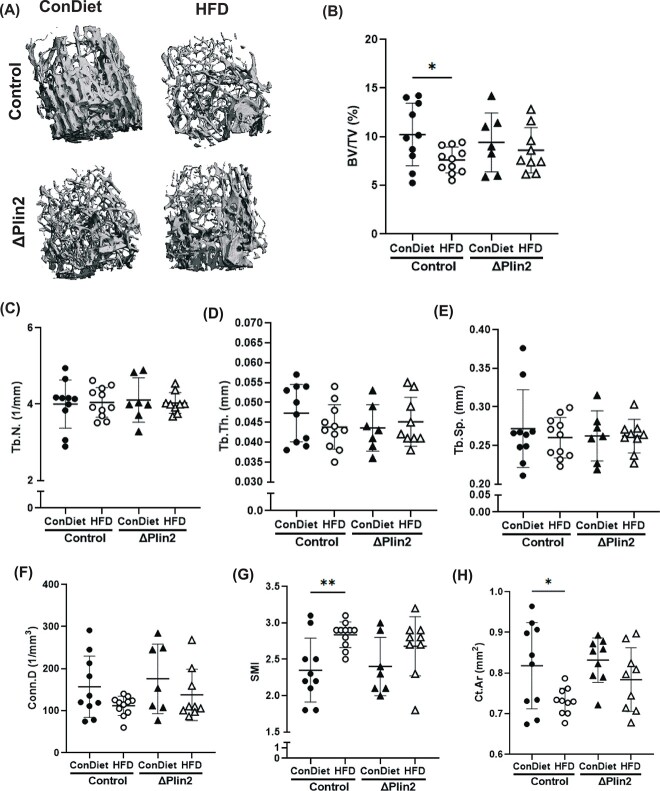
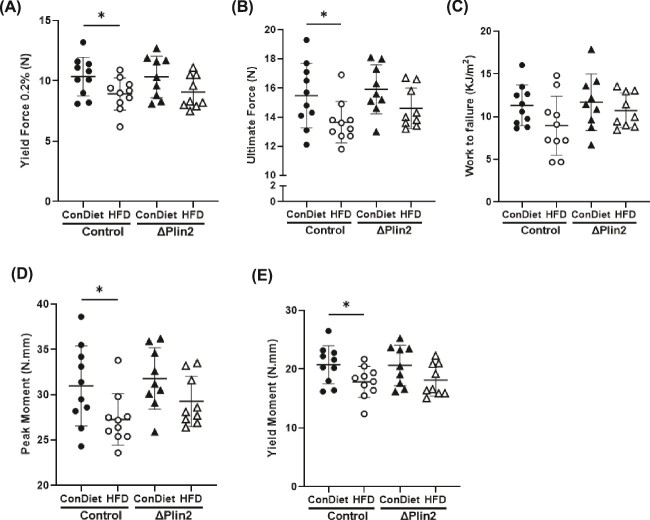
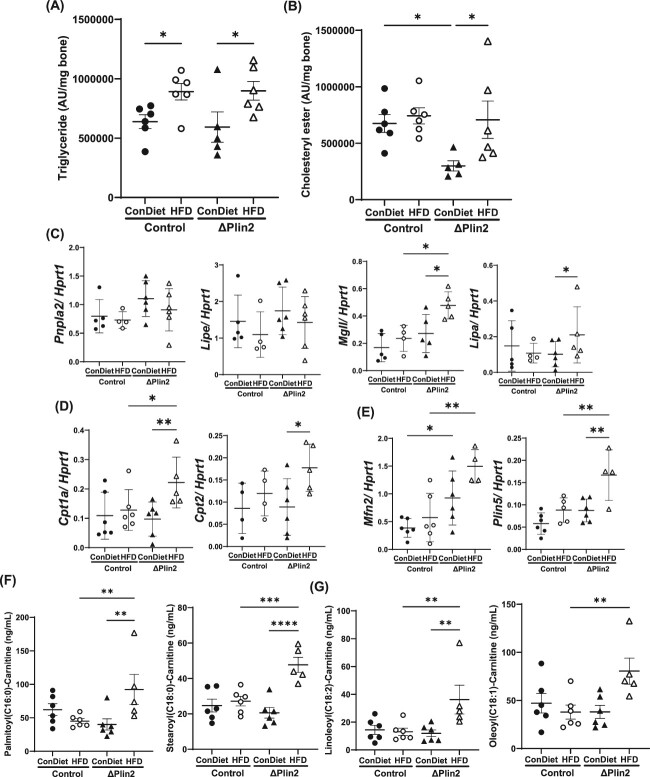
Bone homeostasis within the skeletal system is predominantly maintained by bone formation and resorption, where formation of new bone involves maturation of stromal cells to mineral and matrix secreting mature osteoblasts, which requires cellular energy or adenosine triphosphate. Alterations in systemic metabolism can influence osteoblast function. In line with this, type 2 diabetes mellitus (T2DM), a common metabolic disorder is also associated with reduced bone formation and increased risk of fracture. Impairment in lipid metabolism is one of the key features associated with T2DM-related pathologies in multiple tissues. Therefore, we tested the hypothesis that the reduced bone formation reported in obese murine models of impaired glucose tolerance is a function of disrupted lipid metabolism in osteoblasts. We first confirmed that mice fed a high-fat diet (HFD) have reduced bone microarchitecture along with lower bone formation rates. Interestingly, osteoblasts from obese mice harbor higher numbers of cytosolic lipid droplets along with decreased bioenergetic profiles compared to control cells. Further supporting this observation, bone cortex demonstrated higher total lipid content in HFD fed mice compared to control-fed mice. As a further proof of principle, we generated a novel murine model to conditionally delete Plin2 in osteoblast-progenitor cells using Prrx1-Cre, to enhance lipid droplet breakdown. Our data demonstrate that knocking down Plin2 in an osteoprogenitor specific manner protects from HFD induced osteoblast dysfunction. Furthermore, the mechanism of action involves enhanced osteoblast fatty acid oxidation. In conclusion, the current studies establish that HFD induced glucose intolerance leads to perturbations in osteoblast lipid metabolism, thus causing lower bone formation, which can be protected against by increasing fatty acid oxidation.
Keywords: diabetes; fracture; lipids; metabolism; perilipin 2.
Plain language summary
Obesity is a chronic condition that results from excess fat, which can result in diseases such as type 2 diabetes mellitus. A striking consequence of diabetes is weaker bones leading to the increased risk of fracture; however, factors contributing to this remain unknown. Due to the connection between altered fat metabolism during diabetes, along with weaker bones, we sought to investigate how bone cell fat metabolism was altered during diabetes. First, we demonstrate that high-fat diet mouse models of diabetes resulted in lower bone formation as a function of fat accumulation in the bone, with lower energy production. We next used a genetic mouse model to promote fat metabolism in bone cells responsible for bone formation. Interestingly, these mice were protected from diabetes-associated bone loss. We were able to demonstrate this protection was through changes in bone cell fat metabolism. Collectively, these data establish that bone cell fat metabolism is critical for bone formation and bone quality, and that conditions such as diabetes result in altered fat metabolism and contribute to bone fragility.
© The Author(s) 2024. Published by Oxford University Press on behalf of the American Society for Bone and Mineral Research.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
