

In planta ectopic expression of two subtypes of tomato cellulose synthase-like M genes affects cell wall integrity and supports a role in arabinogalactan and/or rhamnogalacturonan-I biosynthesis
- PMID: 39658008
- PMCID: PMC11775392
- DOI: 10.1093/pcp/pcae145
In planta ectopic expression of two subtypes of tomato cellulose synthase-like M genes affects cell wall integrity and supports a role in arabinogalactan and/or rhamnogalacturonan-I biosynthesis
Abstract
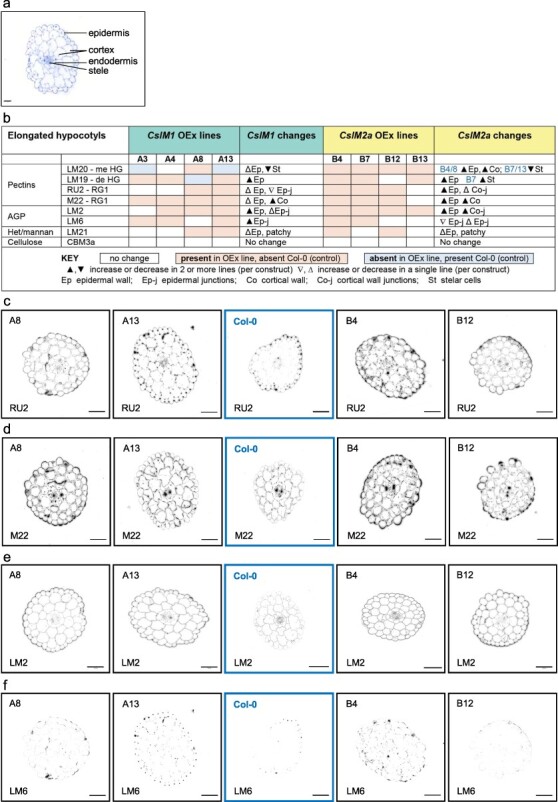
Diversification of the cellulose synthase superfamily of glycosyltransferases has provided plants with the ability to synthesize varied cell wall polysaccharides such as xyloglucan, mannans, and the mixed-linkage glucans of cereals. Surprisingly, some but not all members of the cellulose synthase-like M (CslM) gene family have recently been shown to be involved in the glycosylation of the aglycone core of a range of triterpenoid saponins. However, no cell wall activity has yet been attributed to any of the CslM gene family members. Here, evolution of the CslM gene family in eudicots is explored to better understand the differences between the two metabolically distinct classes of CslMs (CslM1 and CslM2) and the very closely related CslGs. To achieve this, a robust tBLASTn approach was developed to identify CslM1, CslM2, and CslG sequences using diagnostic peptides, suitable for complex genomes using unannotated and short-read datasets. To ascertain whether both CslM1 and CslM2 proteins have cell wall functions, in addition to the 'saponin' role of CslM2, tomato CslM1 and CslM2 genes were ectopically expressed in Arabidopsis thaliana by stable transformation and in the transient Nicotiana benthamiana system. Transformed plants were analysed with immunofluorescence, immunogold transmission electron microscopy, and cell wall polysaccharides were extracted for monosaccharide linkage analysis. Our results support a role for both CslM1 and CslM2 in the biosynthesis of type II arabinogalactan linkages, generating new insight into how the diverse functions of CslMs can coexist and providing clear targets for future research.
Keywords: Solanum lycopersicum; cellulose synthase-like; eudicots; heterologous gene expression; immunofluorescence; plant cell walls.
© The Author(s) 2025. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.
Conflict of interest statement
None declared.
Figures






References
-
- Aharoni A., Sonawane P.D., Itkin M. and Jozwiak A. (2020) Cellulose-synthase-like enzymes and uses thereof. World Patent WO2020049572A1.
-
- APG IV . (2016) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 181: 1–20.
-
- Atmodjo M.A., Hao Z.Y. and Mohnen D. (2013) Evolving views of pectin biosynthesis. Annu. Rev. Plant Biol. 64: 747–779. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
