

Rc3h1 negatively regulates osteoclastogenesis by limiting energy metabolism
- PMID: 39659568
- PMCID: PMC11626950
- DOI: 10.7150/thno.99565
Rc3h1 negatively regulates osteoclastogenesis by limiting energy metabolism
Abstract
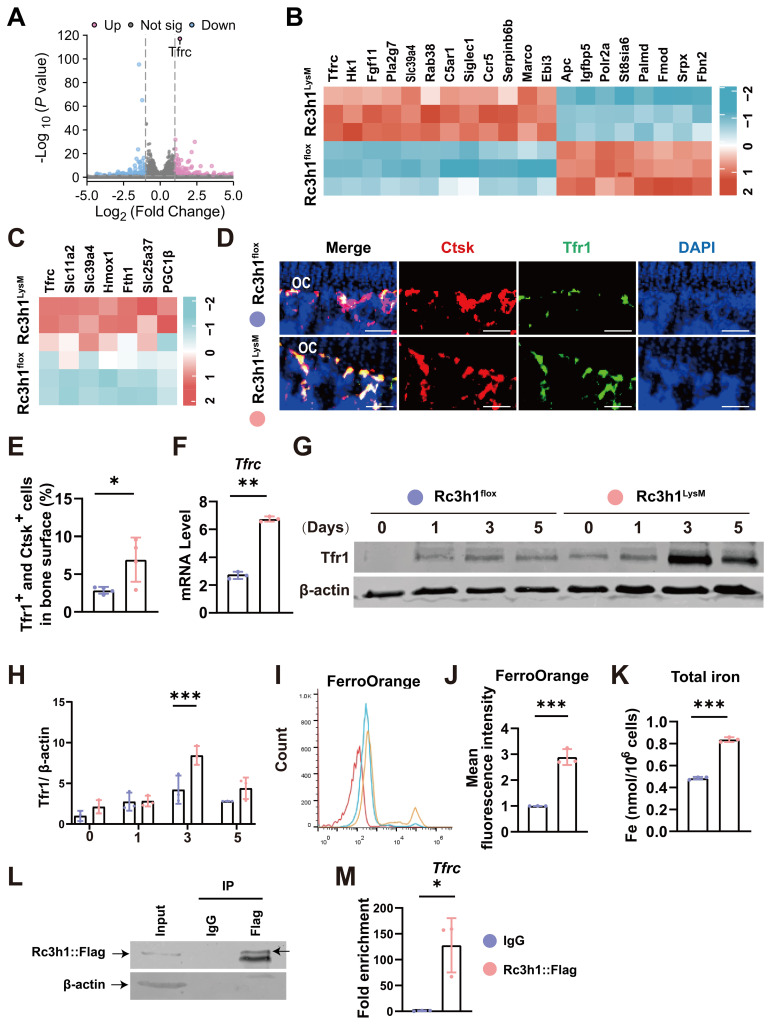
Rationale: Osteoclasts are giant bone-resorbing cells that need vigorous mitochondrial respiration to support their activation. Rc3h1, an RNA-binding protein, precisely governs the homeostasis of mRNA. However, the precise role of Rc3h1 in regulating iron metabolism and mitochondrial respiration in osteoclasts is not yet understood. Methods: We generated Rc3h1-deficient mice in osteoclast precursors and mature osteoclasts. The bone mass and osteoclast activity in bone tissues were evaluated. Moreover, we assessed the differentiation, bone resorption, iron content, and mitochondrial function of osteoclasts in vitro. In the end, the target gene of Rc3h1 and its role in mediating the effect of Rc3h1 on mitochondrial respiration in osteoclasts were further investigated. Results: Mice lacking Rc3h1 exhibit low bone mass. In addition, Rc3h1 deletion in osteoclasts significantly promotes osteoclast activation. Mechanistically, Rc3h1 post-transcriptionally represses the expression of transferrin receptor 1 (Tfr1), restricting iron absorption and mitochondrial respiration in osteoclasts. Inhibition of Tfr1 in Rc3h1-deficient osteoclasts diminishes excessive osteoclast formation and mitochondrial respiration. Conclusion: These findings suggest that Rc3h1 has a negative effect on osteoclast activation via limiting iron resorption and mitochondrial respiration. Finally, targeting the Rc3h1/Tfr1 axis might represent a potential therapeutic approach for bone-loss diseases.
Keywords: Rc3h1; Tfr1; mitochondria; osteoclast; osteoporosis.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures





References
-
- Lemma S, Sboarina M, Porporato PE, Zini N, Sonveaux P, Di Pompo G. et al. Energy metabolism in osteoclast formation and activity. Int J Biochem Cell Biol. 2016;79:168–80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
