

Urolithin A and nicotinamide riboside differentially regulate innate immune defenses and metabolism in human microglial cells
- PMID: 39665042
- PMCID: PMC11631940
- DOI: 10.3389/fnagi.2024.1503336
Urolithin A and nicotinamide riboside differentially regulate innate immune defenses and metabolism in human microglial cells
Abstract
Introduction: During aging, many cellular processes, such as autophagic clearance, DNA repair, mitochondrial health, metabolism, nicotinamide adenine dinucleotide (NAD+) levels, and immunological responses, become compromised. Urolithin A (UA) and Nicotinamide Riboside (NR) are two naturally occurring compounds known for their anti-inflammatory and mitochondrial protective properties, yet the effects of these natural substances on microglia cells have not been thoroughly investigated. As both UA and NR are considered safe dietary supplements, it is equally important to understand their function in normal cells and in disease states.
Methods: This study investigates the effects of UA and NR on immune signaling, mitochondrial function, and microglial activity in a human microglial cell line (HMC3).
Results: Both UA and NR were shown to reduce DNA damage-induced cellular senescence. However, they differentially regulated gene expression related to neuroinflammation, with UA enhancing cGAS-STING pathway activation and NR displaying broader anti-inflammatory effects. Furthermore, UA and NR differently influenced mitochondrial dynamics, with both compounds improving mitochondrial respiration but exhibiting distinct effects on production of reactive oxygen species and glycolytic function.
Discussion: These findings underscore the potential of UA and NR as therapeutic agents in managing neuroinflammation and mitochondrial dysfunction in neurodegenerative diseases.
Keywords: aging; innate immune signaling; microglia; mitochondrial health; nicotinamide riboside; urolithin A.
Copyright © 2024 Madsen, Navarro, Gasparini, Park, Li, Croteau and Bohr.
Conflict of interest statement
VB had a CRADA with ChromaDex Inc and they provided NR for these studies. VB is a board member of ChromaDex Inc, a provider of nicotinamide riboside. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The handling editor KF declared a shared affiliation with the author(s) HM, CN, EG, ZL, VB at the time of review.
Figures
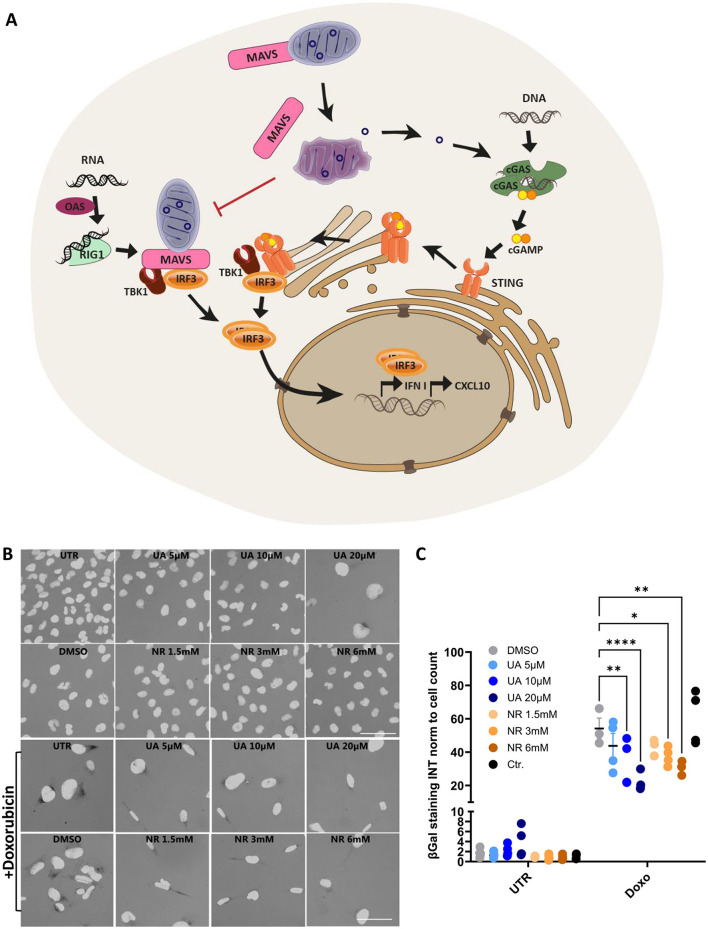
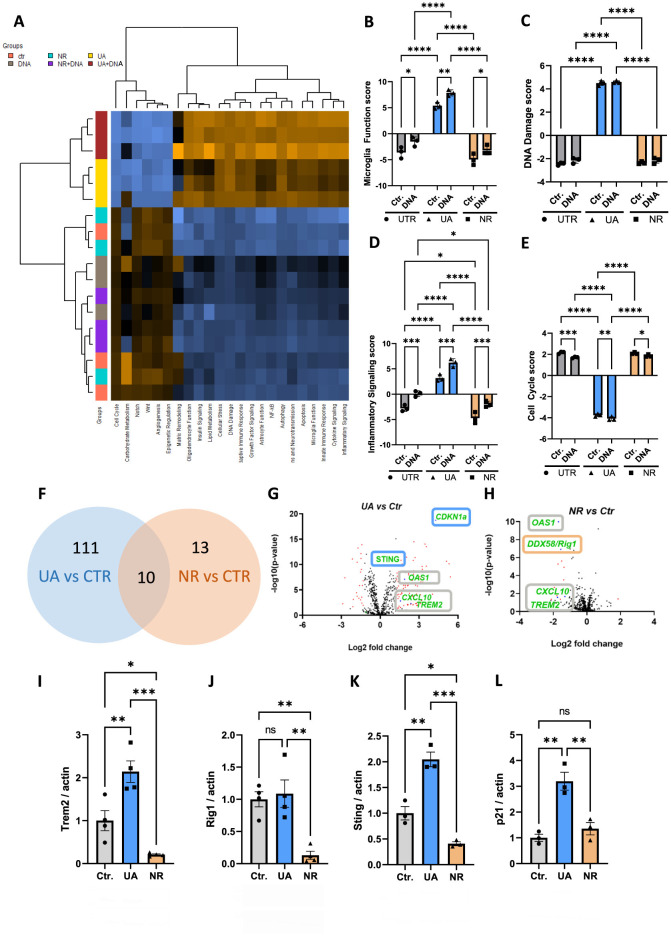
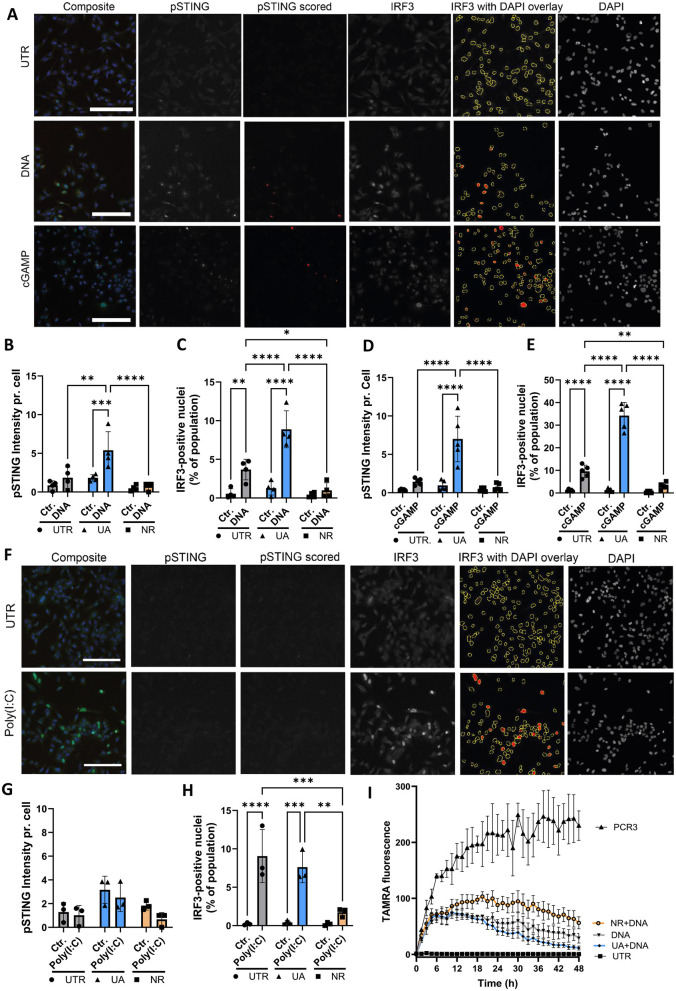
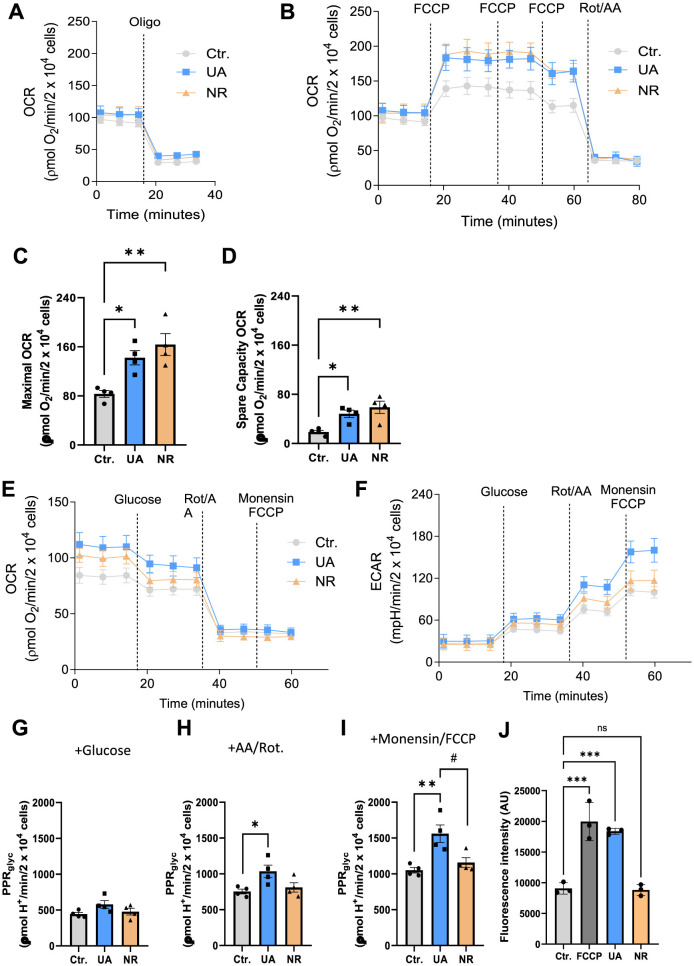
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials