

A synthetic protein-level neural network in mammalian cells
- PMID: 39666795
- PMCID: PMC11758091
- DOI: 10.1126/science.add8468
A synthetic protein-level neural network in mammalian cells
Abstract
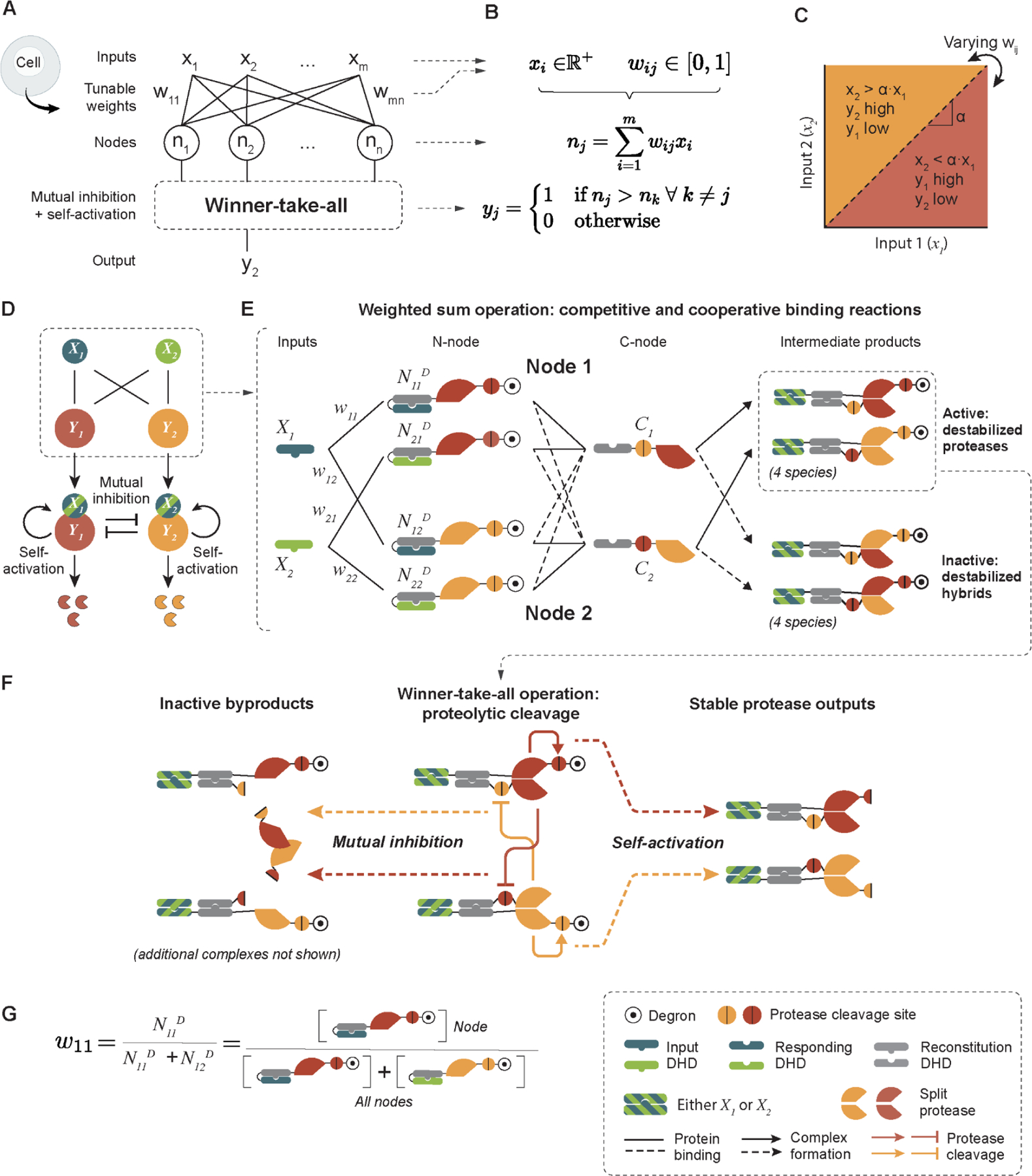
Artificial neural networks provide a powerful paradigm for nonbiological information processing. To understand whether similar principles could enable computation within living cells, we combined de novo-designed protein heterodimers and engineered viral proteases to implement a synthetic protein circuit that performs winner-take-all neural network classification. This "perceptein" circuit combines weighted input summation through reversible binding interactions with self-activation and mutual inhibition through irreversible proteolytic cleavage. These interactions collectively generate a large repertoire of distinct protein species stemming from up to eight coexpressed starting protein species. The complete system achieves multi-output signal classification with tunable decision boundaries in mammalian cells and can be used to conditionally control cell death. These results demonstrate how engineered protein-based networks can enable programmable signal classification in living cells.
Conflict of interest statement
Competing interests: Z.C. and M.B.E. have filed a provisional patent application based on this work. M.B.E. is a co-founder, scientific advisory board member, or consultant at TeraCyte, Primordium Labs (now Plasmidsaurus), and Spatial Genomics.
Figures



Comment in
-
Bringing neural networks to life.Science. 2024 Dec 13;386(6727):1225-1226. doi: 10.1126/science.adu1327. Epub 2024 Dec 12. Science. 2024. PMID: 39666818
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
