

Membrane potential states gate synaptic consolidation in human neocortical tissue
- PMID: 39668146
- PMCID: PMC11638263
- DOI: 10.1038/s41467-024-53901-2
Membrane potential states gate synaptic consolidation in human neocortical tissue
Abstract
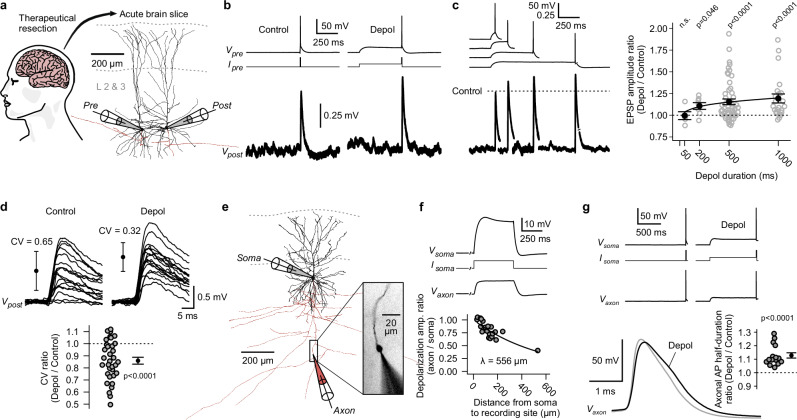
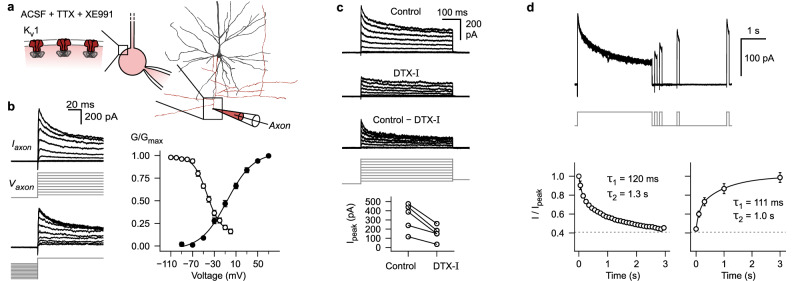
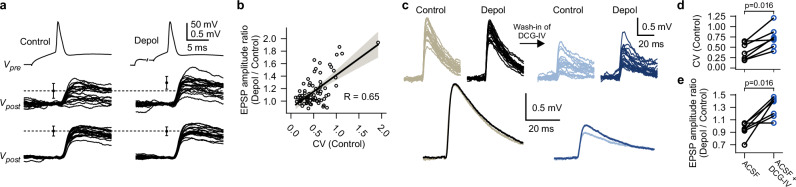
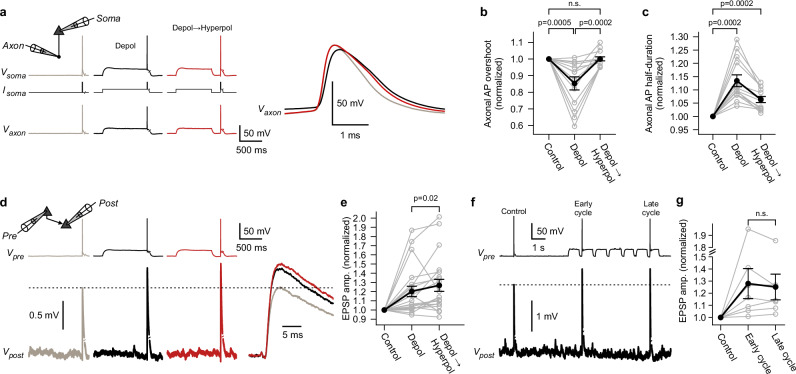
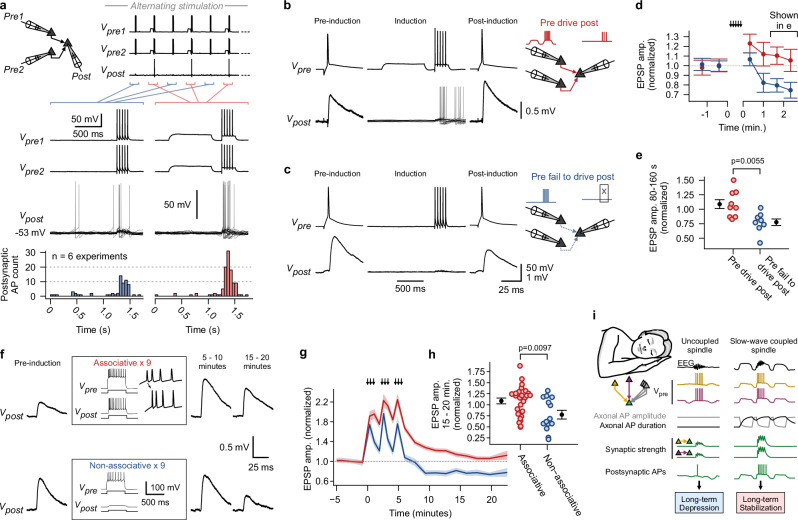
Synaptic mechanisms that contribute to human memory consolidation remain largely unexplored. Consolidation critically relies on sleep. During slow wave sleep, neurons exhibit characteristic membrane potential oscillations known as UP and DOWN states. Coupling of memory reactivation to these slow oscillations promotes consolidation, though the underlying mechanisms remain elusive. Here, we performed axonal and multineuron patch-clamp recordings in acute human brain slices, obtained from neurosurgeries, to show that sleep-like UP and DOWN states modulate axonal action potentials and temporarily enhance synaptic transmission between neocortical pyramidal neurons. Synaptic enhancement by UP and DOWN state sequences facilitates recruitment of postsynaptic action potentials, which in turn results in long-term stabilization of synaptic strength. In contrast, synapses undergo lasting depression if presynaptic neurons fail to recruit postsynaptic action potentials. Our study offers a mechanistic explanation for how coupling of neural activity to slow waves can cause synaptic consolidation, with potential implications for brain stimulation strategies targeting memory performance.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
-
- Klinzing, J. G., Niethard, N. & Born, J. Mechanisms of systems memory consolidation during sleep. Nat. Neurosci.22, 1598–1610 (2019). - PubMed
-
- Corkin, S. What’s new with the amnesic patient H.M.? Nat. Rev. Neurosci.3, 153–160 (2002). - PubMed
-
- Frankland, P. W. & Bontempi, B. The organization of recent and remote memories. Nat. Rev. Neurosci.6, 119–130 (2005). - PubMed
-
- Buzsaki, G. The hippocampo-neocortical dialogue. Cereb. Cortex6, 81–92 (1996). - PubMed
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
