Emergent actin flows explain distinct modes of gliding motility
- PMID: 39669527
- PMCID: PMC11631758
- DOI: 10.1038/s41567-024-02652-4
Emergent actin flows explain distinct modes of gliding motility
Abstract
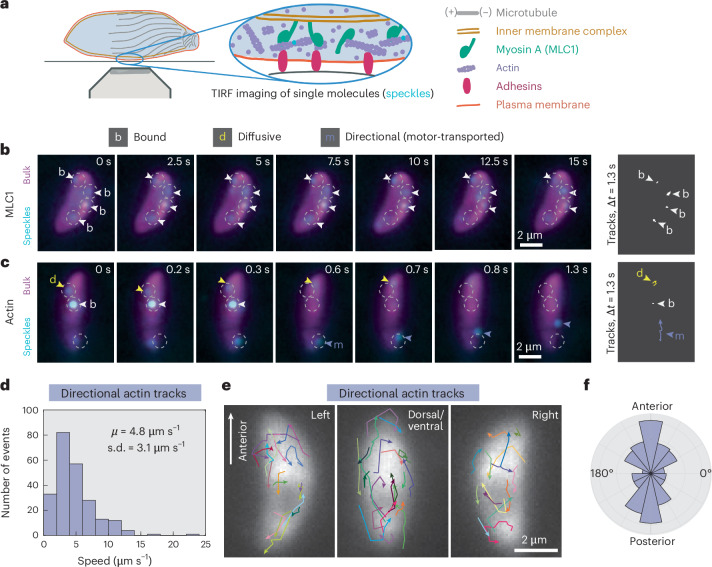
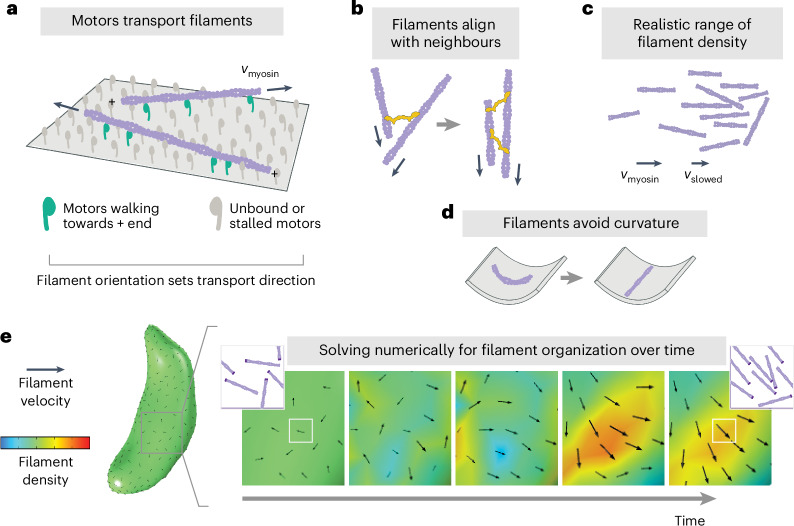
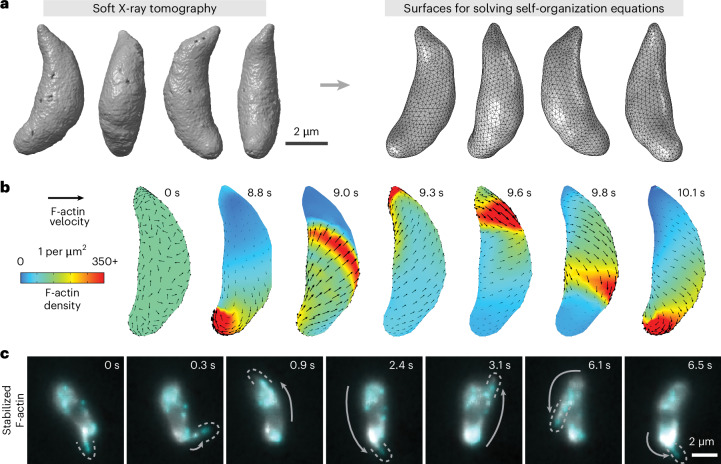
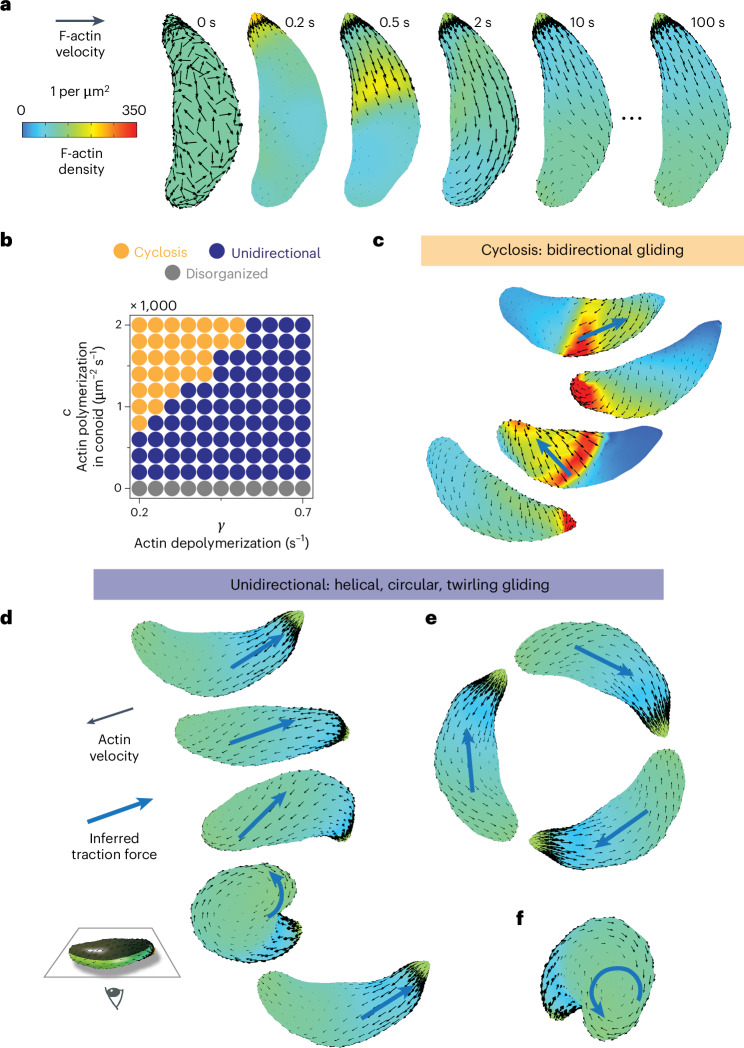
During host infection, Toxoplasma gondii and related unicellular parasites move using gliding, which differs fundamentally from other known mechanisms of eukaryotic cell motility. Gliding is thought to be powered by a thin layer of flowing filamentous (F)-actin sandwiched between the plasma membrane and a myosin-covered inner membrane complex. How this surface actin layer drives the various gliding modes observed in experiments-helical, circular, twirling and patch, pendulum or rolling-is unclear. Here we suggest that F-actin flows arise through self-organization and develop a continuum model of emergent F-actin flow within the confines provided by Toxoplasma geometry. In the presence of F-actin turnover, our model predicts the emergence of a steady-state mode in which actin transport is largely directed rearward. Removing F-actin turnover leads to actin patches that recirculate up and down the cell, which we observe experimentally for drug-stabilized actin bundles in live Toxoplasma gondii parasites. These distinct self-organized actin states can account for observed gliding modes, illustrating how different forms of gliding motility can emerge as an intrinsic consequence of the self-organizing properties of F-actin flow in a confined geometry.
Keywords: Biological physics; Cellular motility.
© The Author(s) 2024.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures