

Temperature-dependent fold-switching mechanism of the circadian clock protein KaiB
- PMID: 39671178
- PMCID: PMC11665890
- DOI: 10.1073/pnas.2412327121
Temperature-dependent fold-switching mechanism of the circadian clock protein KaiB
Abstract
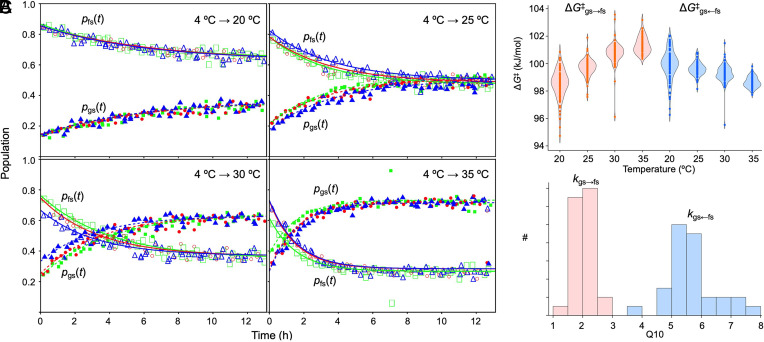
The oscillator of the cyanobacterial circadian clock relies on the ability of the KaiB protein to switch reversibly between a stable ground-state fold (gsKaiB) and an unstable fold-switched fold (fsKaiB). Rare fold-switching events by KaiB provide a critical delay in the negative feedback loop of this posttranslational oscillator. In this study, we experimentally and computationally investigate the temperature dependence of fold switching and its mechanism. We demonstrate that the stability of gsKaiB increases with temperature compared to fsKaiB and that the Q10 value for the gsKaiB → fsKaiB transition is nearly three times smaller than that for the reverse transition in a construct optimized for NMR studies. Simulations and native-state hydrogen-deuterium exchange NMR experiments suggest that fold switching can involve both partially and completely unfolded intermediates. The simulations predict that the transition state for fold switching coincides with isomerization of conserved prolines in the most rapidly exchanging region, and we confirm experimentally that proline isomerization is a rate-limiting step for fold switching. We explore the implications of our results for temperature compensation, a hallmark of circadian clocks, through a kinetic model.
Keywords: NMR; circadian clock; molecular dynamics; protein folding; temperature compensation.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures






Update of
-
Temperature-Dependent Fold-Switching Mechanism of the Circadian Clock Protein KaiB.bioRxiv [Preprint]. 2024 May 21:2024.05.21.594594. doi: 10.1101/2024.05.21.594594. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Dec 17;121(51):e2412327121. doi: 10.1073/pnas.2412327121. PMID: 38826295 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
