

Integrin adhesome axis inhibits the RPM-1 ubiquitin ligase signaling hub to regulate growth cone and axon development
- PMID: 39671436
- PMCID: PMC11642917
- DOI: 10.1371/journal.pgen.1011496
Integrin adhesome axis inhibits the RPM-1 ubiquitin ligase signaling hub to regulate growth cone and axon development
Abstract
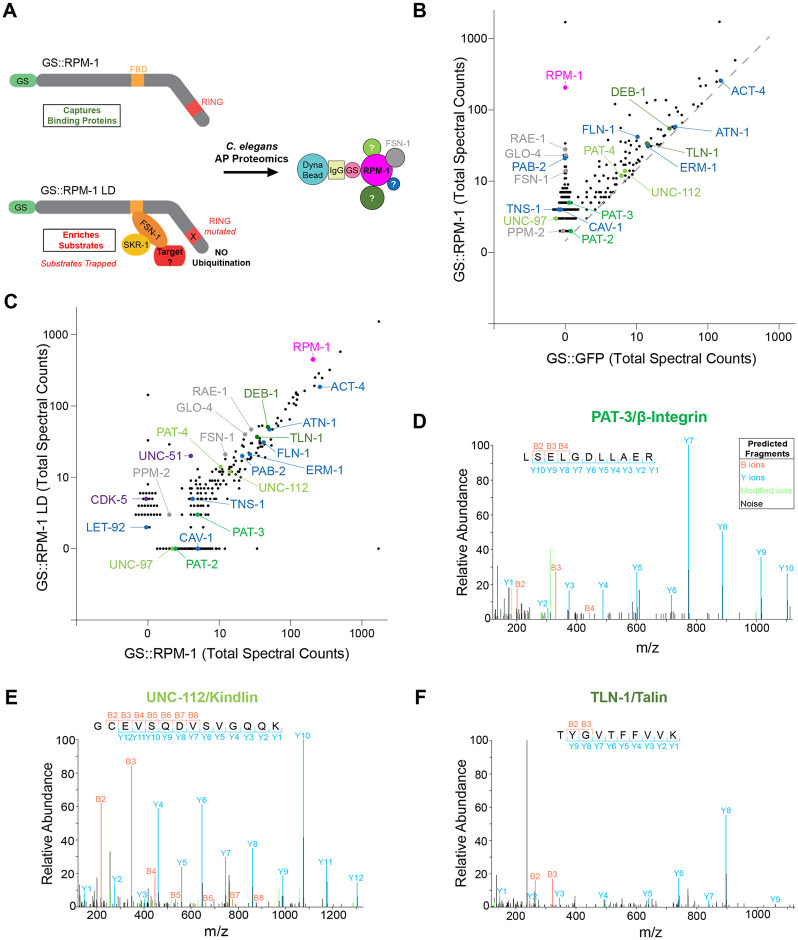
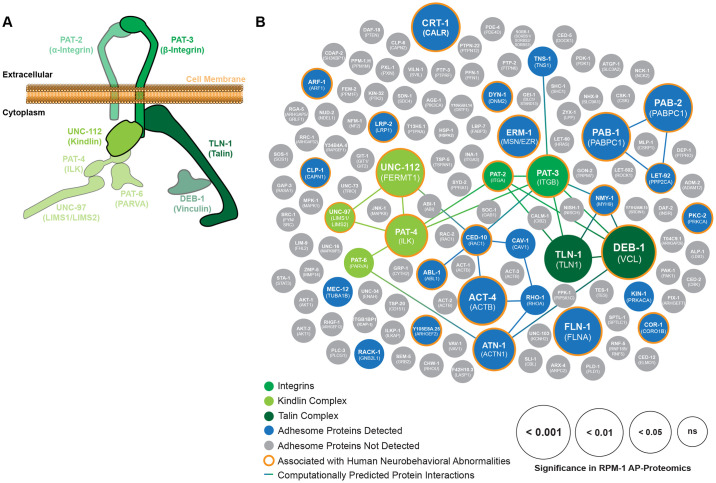
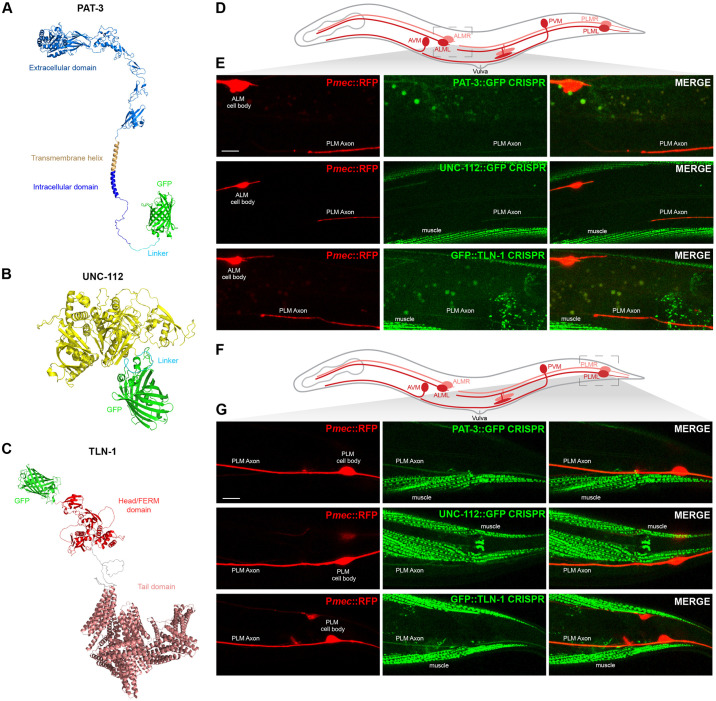
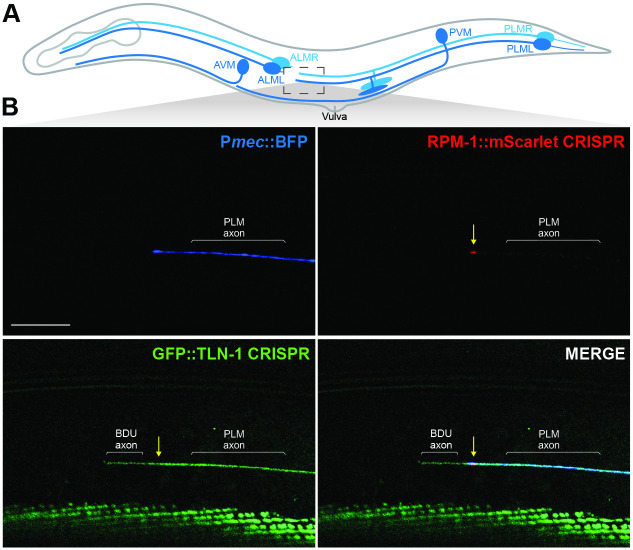
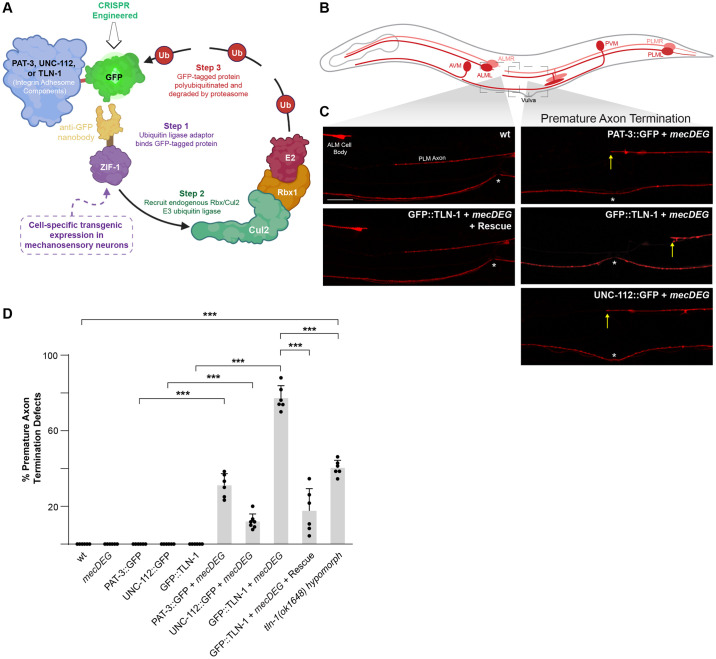
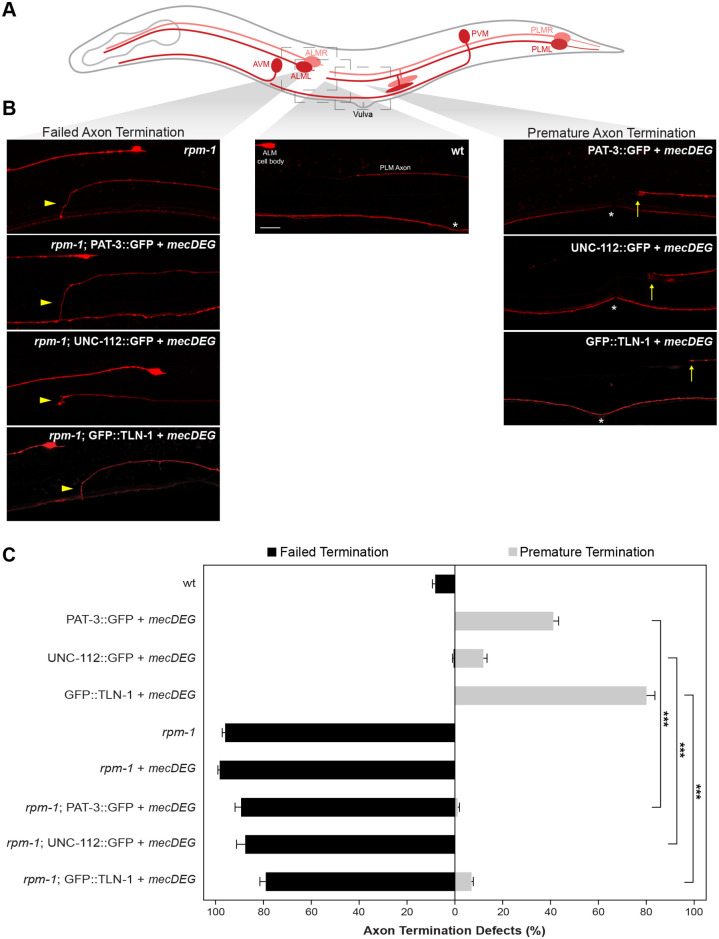
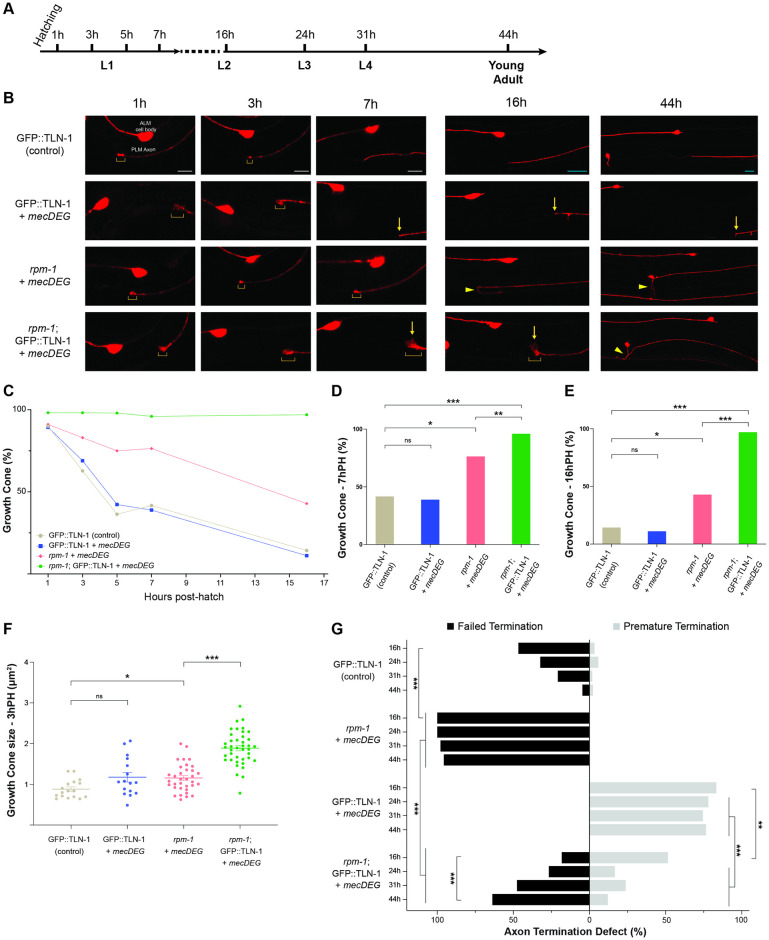
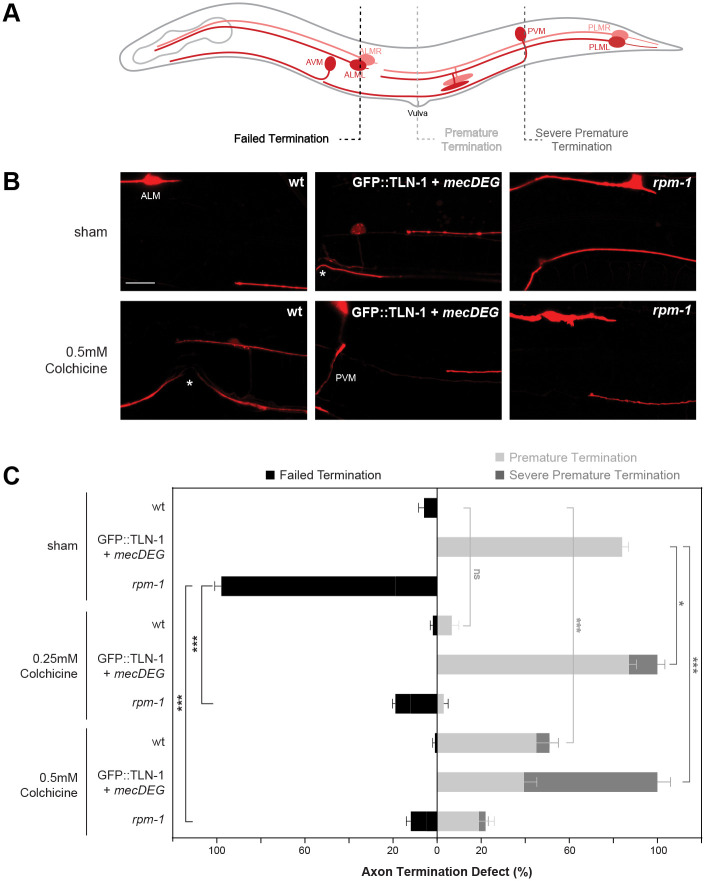
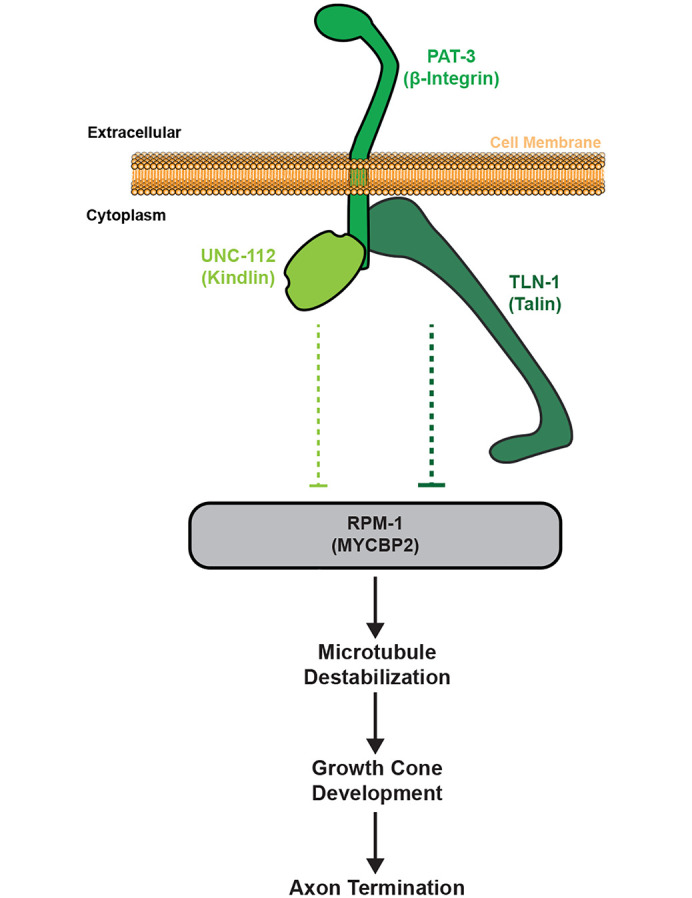
Integrin signaling plays important roles in development and disease. An adhesion signaling network called the integrin adhesome has been principally defined using bioinformatics and cell-based proteomics. To date, the adhesome has not been studied using integrated proteomic and genetic approaches. Here, proteomic studies in C. elegans identified physical associations between the RPM-1 ubiquitin ligase signaling hub and numerous adhesome components including Talin (TLN-1), Kindlin (UNC-112) and β-integrin (PAT-3). C. elegans RPM-1 is orthologous to human MYCBP2, a prominent player in nervous system development recently associated with a neurodevelopmental disorder. After curating and updating the conserved C. elegans adhesome, we identified an adhesome subnetwork physically associated with RPM-1 that has extensive links to human neurobehavioral abnormalities. Using neuron-specific, CRISPR loss-of-function strategies, we demonstrate that a PAT-3/UNC-112/TLN-1 adhesome axis regulates axon termination in mechanosensory neurons by inhibiting RPM-1. Developmental time-course studies and pharmacological results suggest TLN-1 inhibition of RPM-1 affects growth cone collapse and microtubule dynamics during axon outgrowth. These results indicate the PAT-3/UNC-112/TLN-1 adhesome axis restricts RPM-1 signaling to ensure axon outgrowth is terminated in a spatially and temporally accurate manner. Thus, our findings orthogonally validate the adhesome using an organismal setting, identify an adhesome axis that inhibits RPM-1 (MYCBP2), and highlight important new links between the adhesome and brain disorders.
Copyright: © 2024 Amezquita et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Update of
-
Axon development is regulated at genetic and proteomic interfaces between the integrin adhesome and the RPM-1 ubiquitin ligase signaling hub.bioRxiv [Preprint]. 2023 Nov 15:2023.11.15.566604. doi: 10.1101/2023.11.15.566604. bioRxiv. 2023. Update in: PLoS Genet. 2024 Dec 13;20(12):e1011496. doi: 10.1371/journal.pgen.1011496. PMID: 38014183 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
