

An SH3-binding allosteric modulator stabilizes the global conformation of the AML-associated Src-family kinase, Hck
- PMID: 39675702
- PMCID: PMC11786751
- DOI: 10.1016/j.jbc.2024.108088
An SH3-binding allosteric modulator stabilizes the global conformation of the AML-associated Src-family kinase, Hck
Abstract
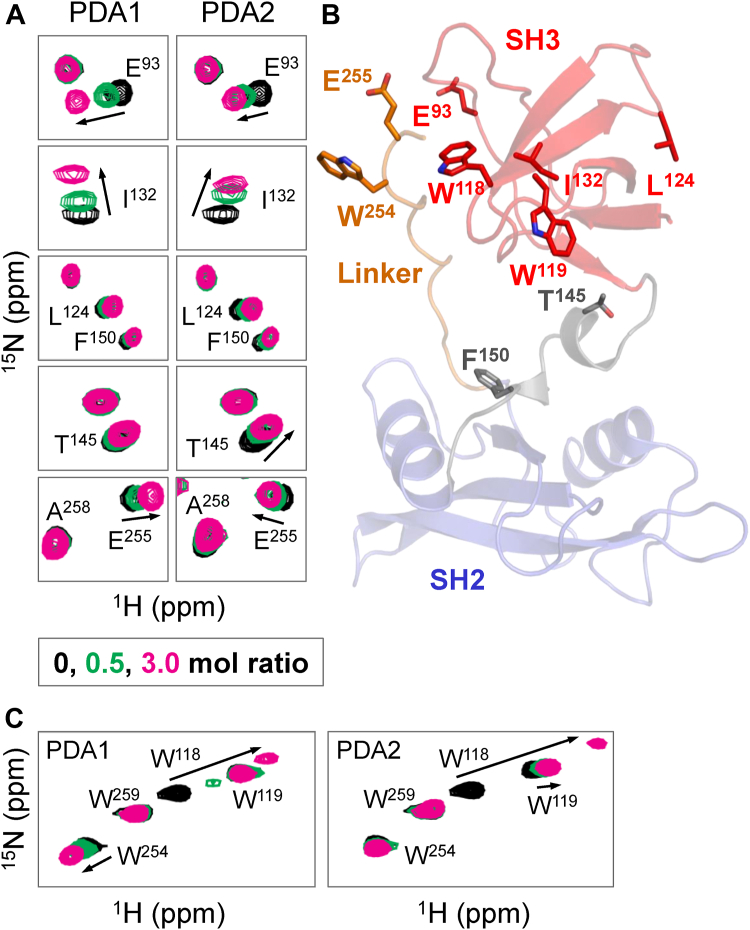
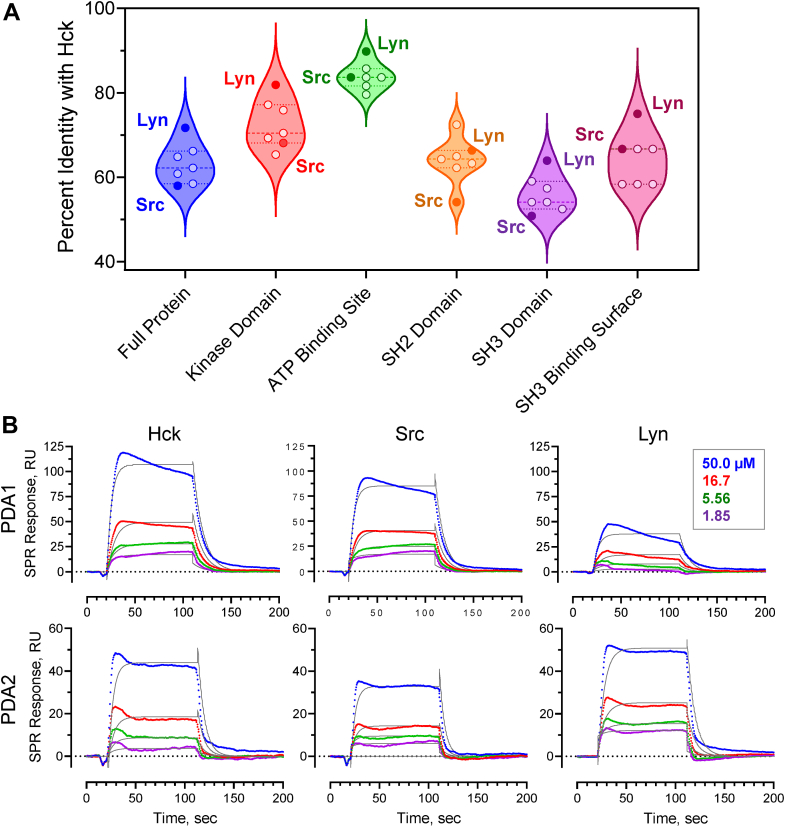
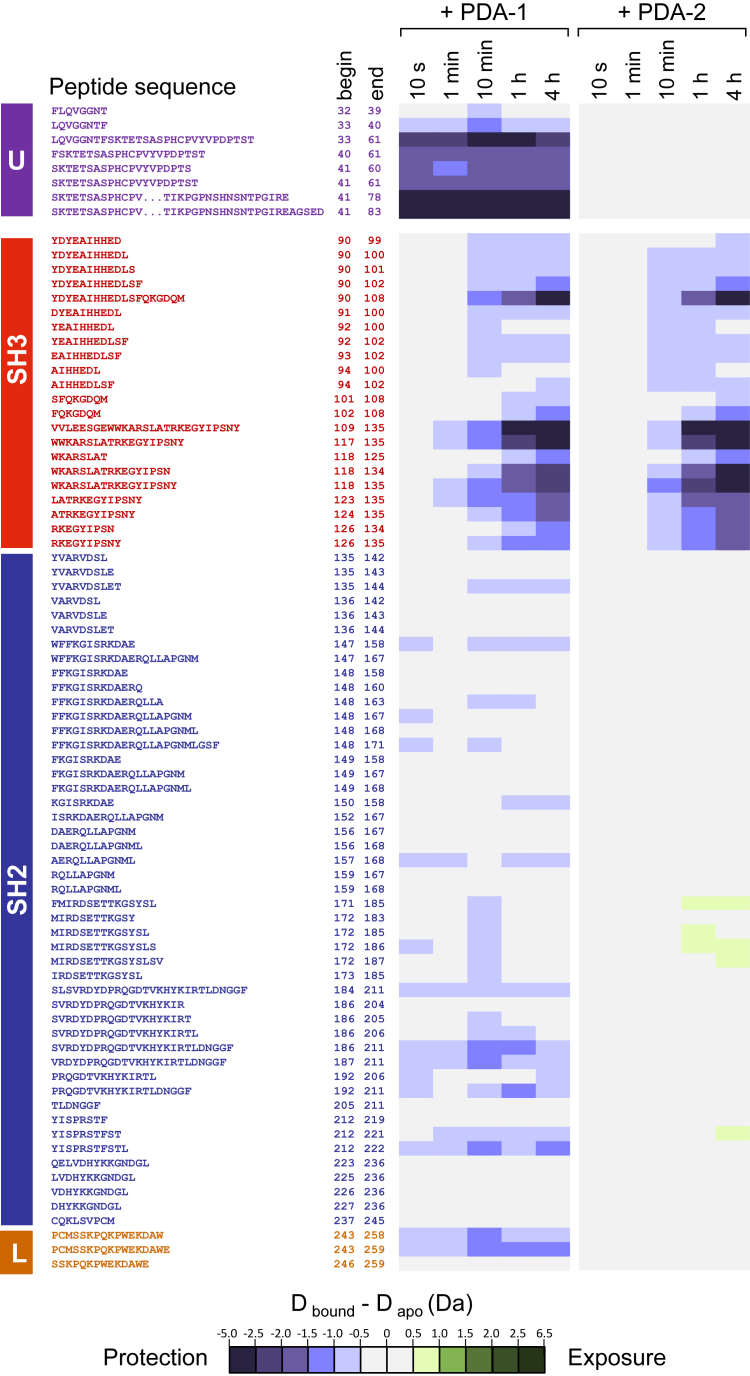
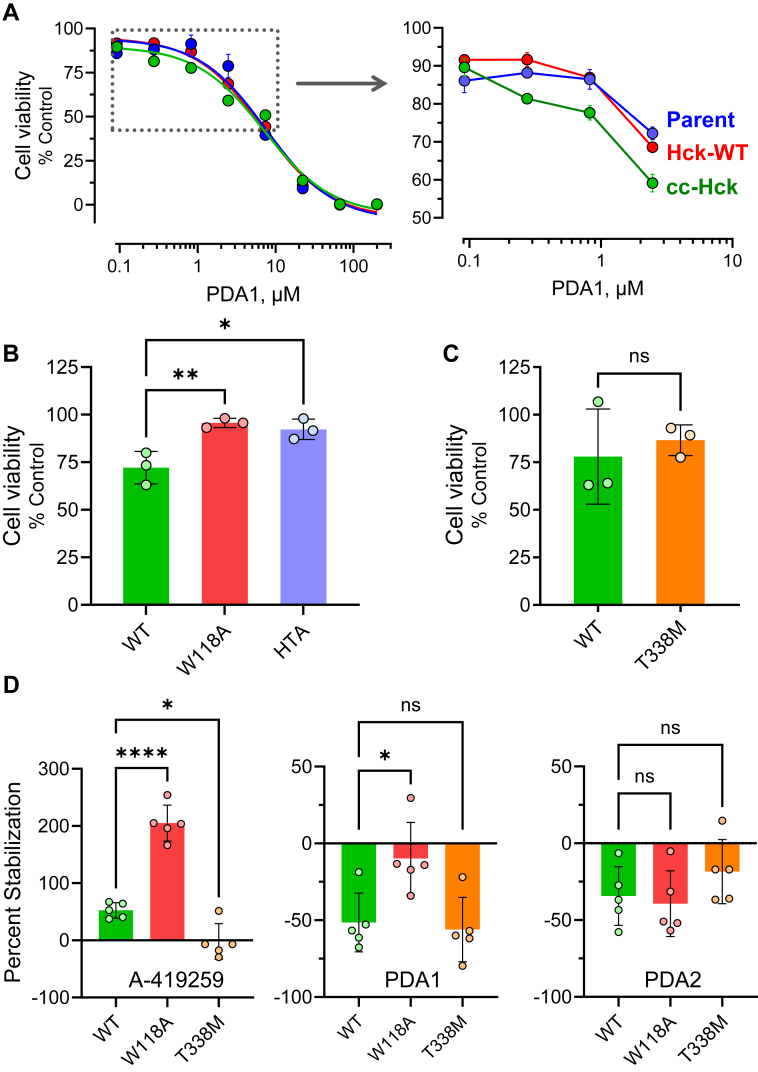
While ATP-site inhibitors for protein-tyrosine kinases are often effective drugs, their clinical utility can be limited by off-target activity and acquired resistance mutations due to the conserved nature of the ATP-binding site. However, combining ATP-site and allosteric kinase inhibitors can overcome these shortcomings in a double-drugging framework. Here we explored the allosteric effects of two pyrimidine diamines, PDA1 and PDA2, on the conformational dynamics and activity of the Src-family tyrosine kinase Hck, a promising drug target for acute myeloid leukemia. Using 1H-15N HSQC NMR, we mapped the binding site for both analogs to the SH3 domain. Despite the shared binding site, PDA1 and PDA2 had opposing effects on near-full-length Hck dynamics by hydrogen-deuterium exchange mass spectrometry, with PDA1 stabilizing and PDA2 disrupting the overall kinase conformation. Kinase activity assays were consistent with these observations, with PDA2 enhancing kinase activity while PDA1 was without effect. Molecular dynamics simulations predicted selective bridging of the kinase domain N-lobe and SH3 domain by PDA1, a mechanism of allosteric stabilization supported by site-directed mutagenesis of N-lobe contact sites. Cellular thermal shift assays confirmed SH3 domain-dependent interaction of PDA1 with WT Hck in myeloid leukemia cells and with a kinase domain gatekeeper mutant (T338M). These results identify PDA1 as a starting point for Src-family kinase allosteric inhibitor development that may work in concert with ATP-site inhibitors to suppress the evolution of resistance.
Keywords: NMR; SH2 domain; SH3 domain; Src-family kinase; hydrogen-deuterium exchange mass spectrometry; kinase inhibitors; tyrosine kinases.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures




References
-
- Boggon T.J., Eck M.J. Structure and regulation of Src family kinases. Oncogene. 2004;23:7918–7927. - PubMed
-
- Chong Y.P., Ia K.K., Mulhern T.D., Cheng H.C. Endogenous and synthetic inhibitors of the Src-family protein tyrosine kinases. Biochim. Biophys. Acta. 2005;1754:210–220. - PubMed
-
- Xu W., Doshi A., Lei M., Eck M.J., Harrison S.C. Crystal structures of c-Src reveal features of its autoinhibitory mechanism. Mol. Cell. 1999;3:629–638. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
