

This is a preprint.
Optogenetically Induced Microtubule Acetylation Unveils the Molecular Dynamics of Actin-Microtubule Crosstalk in Directed Cell Migration
- PMID: 39677776
- PMCID: PMC11642777
- DOI: 10.1101/2024.12.01.626286
Optogenetically Induced Microtubule Acetylation Unveils the Molecular Dynamics of Actin-Microtubule Crosstalk in Directed Cell Migration
Abstract
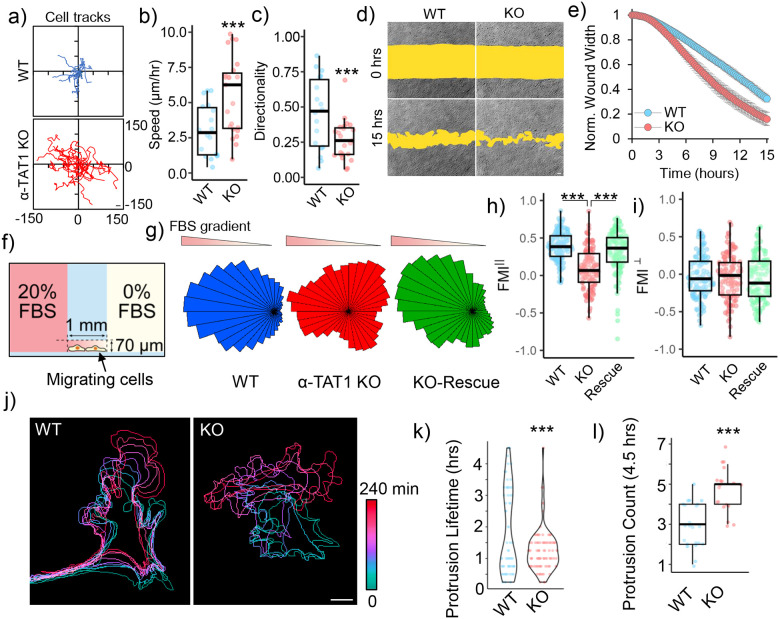
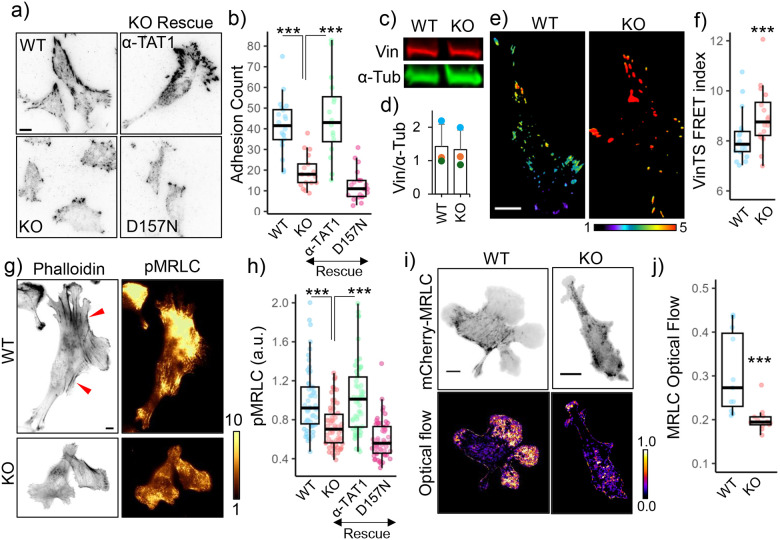
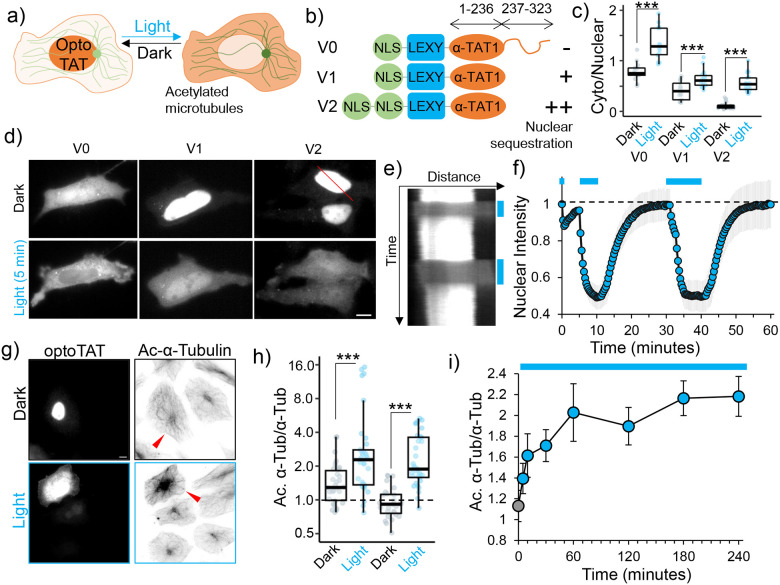
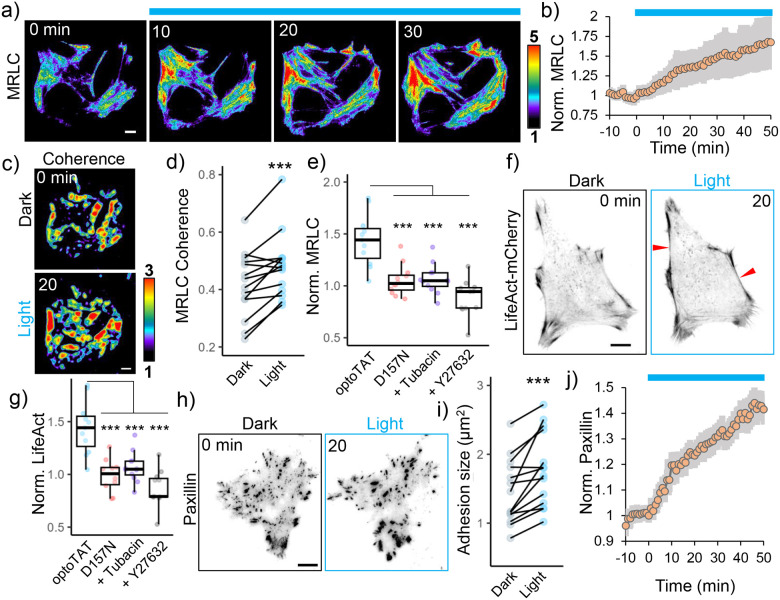
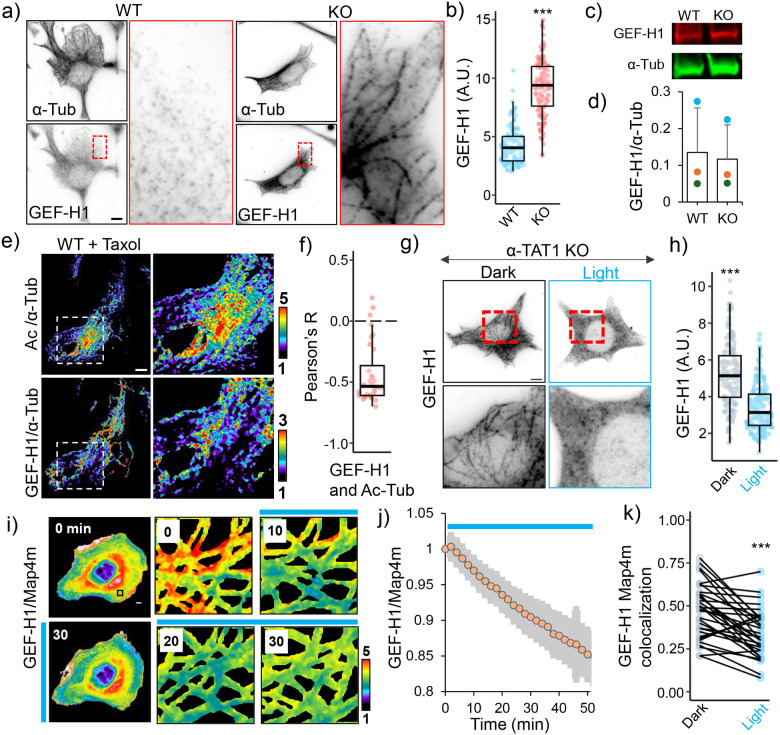
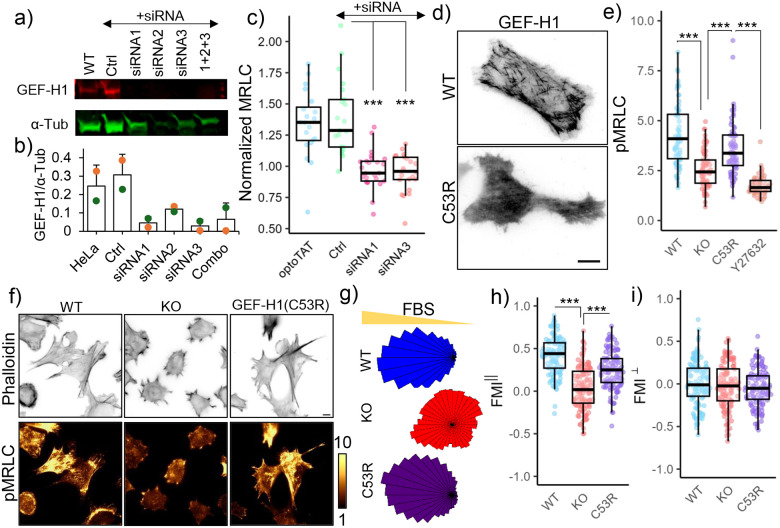
Microtubule acetylation is implicated in regulating cell motility, yet its physiological role in directional migration and the underlying molecular mechanisms have remained unclear. This knowledge gap has persisted primarily due to a lack of tools capable of rapidly manipulating microtubule acetylation in actively migrating cells. To overcome this limitation and elucidate the causal relationship between microtubule acetylation and cell migration, we developed a novel optogenetic actuator, optoTAT, which enables precise and rapid induction of microtubule acetylation within minutes in live cells. Using optoTAT, we observed striking and rapid responses at both molecular and cellular level. First, microtubule acetylation triggers release of the RhoA activator GEF-H1 from sequestration on microtubules. This release subsequently enhances actomyosin contractility and drives focal adhesion maturation. These subcellular processes collectively promote sustained directional cell migration. Our findings position GEF-H1 as a critical molecular responder to microtubule acetylation in the regulation of directed cell migration, revealing a dynamic crosstalk between the actin and microtubule cytoskeletal networks.
Conflict of interest statement
Competing Interests The authors declare that there is a pending patent application related to optoTAT.
Figures
References
-
- Janke C. & Magiera M. M. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. Mol. Cell Biol. 21, 307–326 (2020). - PubMed
-
- L’Hernault S. W. & Rosenbaum J. L. Chlamydomonas alpha-tubulin is posttranslationally modified by acetylation on the epsilon-amino group of a lysine. Biochemistry 24, 473–478 (1985). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous