

Suppressed Protein Translation Caused by MSP-8 Deficiency Determines Fungal Multidrug Resistance with Fitness Cost
- PMID: 39679802
- PMCID: PMC11809369
- DOI: 10.1002/advs.202412514
Suppressed Protein Translation Caused by MSP-8 Deficiency Determines Fungal Multidrug Resistance with Fitness Cost
Abstract
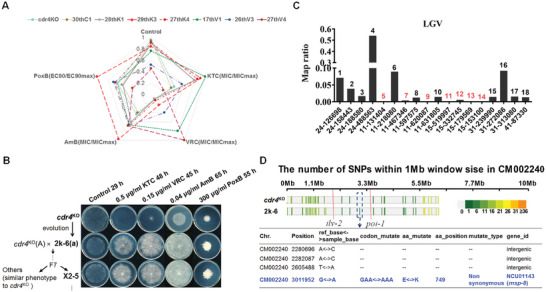
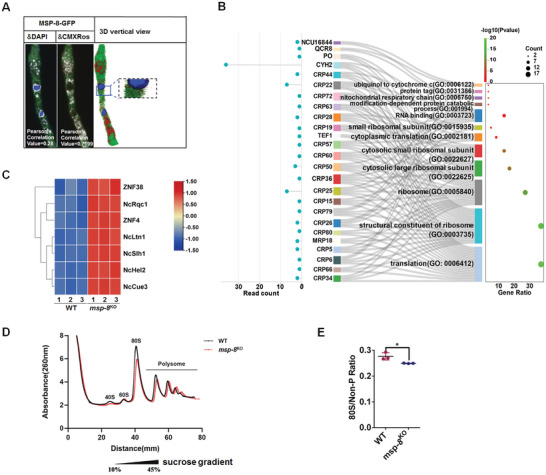
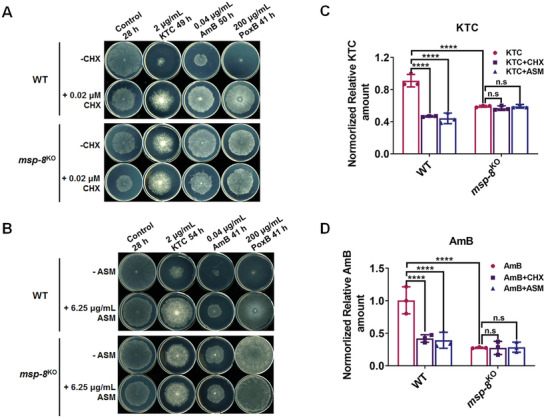
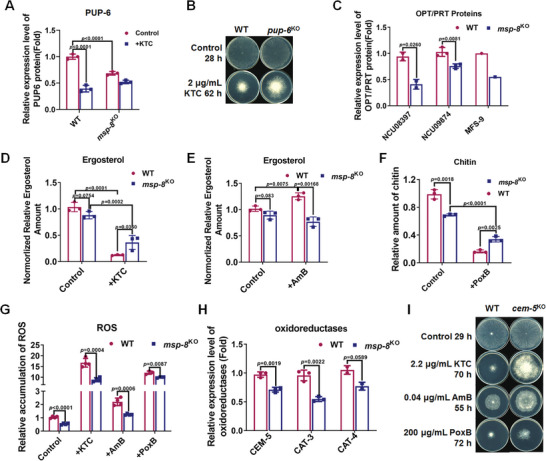
Antifungal resistance, particularly the rise of multidrug-resistance strains, poses a significant public health threat. In this study, the study identifies a novel multidrug-resistance gene, msp-8, encoding a helicase, through experimental evolution with Neurospora crassa as a model. Deletion of msp-8 conferred multidrug resistance in N. crassa, Aspergillus fumigatus, and Fusarium verticillioides. However, the transcript levels of genes encoding known drug targets or efflux pumps remain unaltered with msp-8 deletion. Interestingly, MSP-8 interacted with ribosomal proteins, and this mutant displays compromised ribosomal function, causing translational disturbance. Notably, inhibition of protein translation enhances resistance to azoles, amphotericin B, and polyoxin B. Furthermore, MSP-8 deficiency or inhibition of translation reduces intracellular ketoconazole accumulation and membrane-bound amphotericin B content, directly causing antifungal resistance. Additionaly, MSP-8 deficiency induces cell wall remodeling, and decreases intracellular ROS levels, further contributing to resistance. The findings reveal a novel multidrug resistance mechanism independent of changes in drug target or efflux pump, while MSP-8 deficiency suppresses protein translation, thereby facilitating the development of resistance with fitness cost. This study provides the first evidence that MSP-8 participates in protein translation and that translation suppression can cause multidrug resistance in fungi, offering new insights into resistance mechanisms in clinical and environmental fungal strains.
Keywords: Aspergillus fumigatus; Fusarium verticilioides; Neurospora crassa; antifungal drug; helicase; multidrug resistance; protein translation.
© 2024 The Author(s). Advanced Science published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Denning D. W., Lancet Infect. Dis. 2024, 24, e428.
-
- Fisher M. C., Hawkins N. J., Sanglard D., Gurr S. J., Science 2018, 360, 739. - PubMed
-
- Scheele B. C., Pasmans F., Skerratt L. F., Berger L., Martel A., Beukema W., Acevedo A. A., Burrowes P. A., Carvalho T., Catenazzi A., De la Riva I., Fisher M. C., Flechas S. V., Foster C. N., Frías‐Álvarez P., Garner T. W. J., Gratwicke B., Guayasamin J. M., Hirschfeld M., Kolby J. E., Kosch T. A., La Marca E., Lindenmayer D. B., Lips K. R., Longo A. V., Maneyro R., McDonald C. A., Mendelson J., Palacios‐Rodriguez P., Parra‐Olea G., et al., Science 2019, 363, 1459. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials