

Examining the NEUROG2 lineage and associated gene expression in human cortical organoids
- PMID: 39680368
- PMCID: PMC11829764
- DOI: 10.1242/dev.202703
Examining the NEUROG2 lineage and associated gene expression in human cortical organoids
Abstract
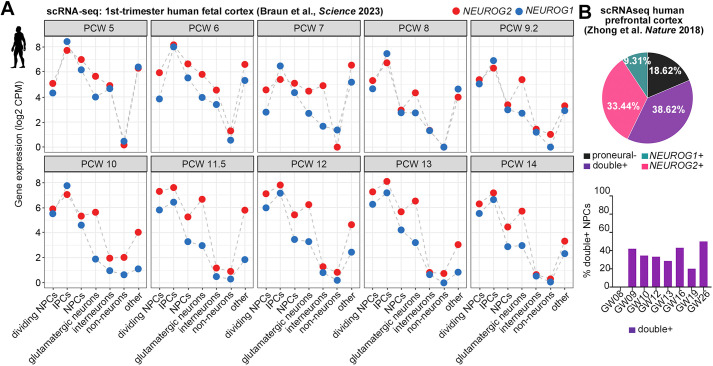
Proneural genes are conserved drivers of neurogenesis across the animal kingdom. How their functions have adapted to guide human-specific neurodevelopmental features is poorly understood. Here, we mined transcriptomic data from human fetal cortices and generated from human embryonic stem cell-derived cortical organoids (COs) to show that NEUROG1 and NEUROG2 are most highly expressed in basal neural progenitor cells, with pseudotime trajectory analyses indicating that NEUROG1-derived lineages predominate early and NEUROG2 lineages later. Using ChIP-qPCR, gene silencing and overexpression studies in COs, we show that NEUROG2 is necessary and sufficient to directly transactivate known target genes (NEUROD1, EOMES, RND2). To identify new targets, we engineered NEUROG2-mCherry knock-in human embryonic stem cells for CO generation. The mCherry-high CO cell transcriptome is enriched in extracellular matrix-associated genes, and two genes associated with human-accelerated regions: PPP1R17 and FZD8. We show that NEUROG2 binds COL1A1, COL3A1 and PPP1R17 regulatory elements, and induces their ectopic expression in COs, although NEUROG2 is not required for this expression. Neurog2 similarly induces Col3a1 and Ppp1r17 in murine P19 cells. These data are consistent with a conservation of NEUROG2 function across mammalian species.
Keywords: NEUROG2; CRISPR gene editing; Cortical organoid; Extracellular matrix; Human-specific features; Lineage tracing; Proneural transcription factor.
© 2025. Published by The Company of Biologists.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures






References
-
- Aydin, B., Kakumanu, A., Rossillo, M., Moreno-Estelles, M., Garipler, G., Ringstad, N., Flames, N., Mahony, S. and Mazzoni, E. O. (2019). Proneural factors Ascl1 and Neurog2 contribute to neuronal subtype identities by establishing distinct chromatin landscapes. Nat. Neurosci. 22, 897-908. 10.1038/s41593-019-0399-y - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
