

Activation of the tick Toll pathway to control infection of Ixodes ricinus by the apicomplexan parasite Babesia microti
- PMID: 39680508
- PMCID: PMC11649134
- DOI: 10.1371/journal.ppat.1012743
Activation of the tick Toll pathway to control infection of Ixodes ricinus by the apicomplexan parasite Babesia microti
Abstract
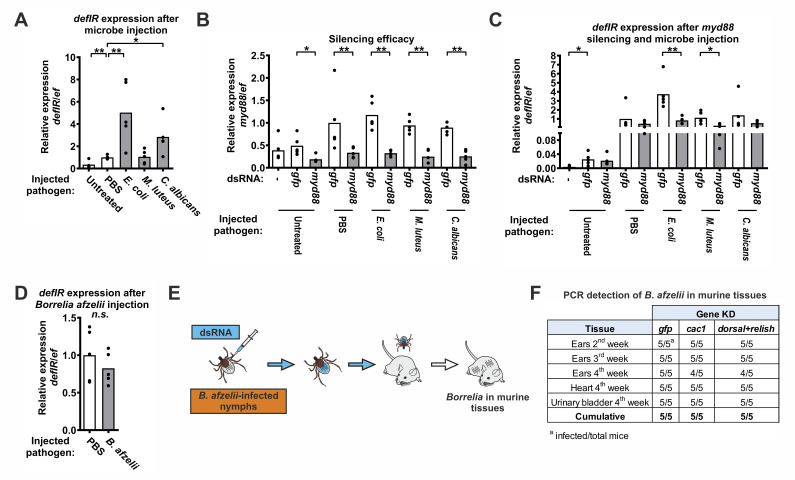
The vector competence of blood-feeding arthropods is influenced by the interaction between pathogens and the immune system of the vector. The Toll and IMD (immune deficiency) signaling pathways play a key role in the regulation of innate immunity in both the Drosophila model and blood-feeding insects. However, in ticks (chelicerates), immune determination for pathogen acquisition and transmission has not yet been fully explored. Here, we have mapped homologs of insect Toll and IMD pathways in the European tick Ixodes ricinus, an important vector of human and animal diseases. We show that most genes of the Toll pathway are well conserved, whereas the IMD pathway has been greatly reduced. We therefore investigated the functions of the individual components of the tick Toll pathway and found that, unlike in Drosophila, it was specifically activated by Gram-negative bacteria. The activation of pathway induced the expression of defensin (defIR), the first identified downstream effector gene of the tick Toll pathway. Borrelia, an atypical bacterium and causative agent of Lyme borreliosis, bypassed Toll-mediated recognition in I. ricinus and also resisted systemic effector molecules when the Toll pathway was activated by silencing its repressor cactus via RNA interference. Babesia, an apicomplexan parasite, also avoided Toll-mediated recognition. Strikingly, unlike Borrelia, the number of Babesia parasites reaching the salivary glands during tick infection was significantly reduced by knocking down cactus. The simultaneous silencing of cactus and dorsal resulted in greater infections and underscored the importance of tick immunity in regulating parasite infections in these important disease vectors.
Copyright: © 2024 Jalovecka et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
