

Amyloid-β oligomers increase the binding and internalization of tau oligomers in human synapses
- PMID: 39688618
- PMCID: PMC11652575
- DOI: 10.1007/s00401-024-02839-2
Amyloid-β oligomers increase the binding and internalization of tau oligomers in human synapses
Abstract
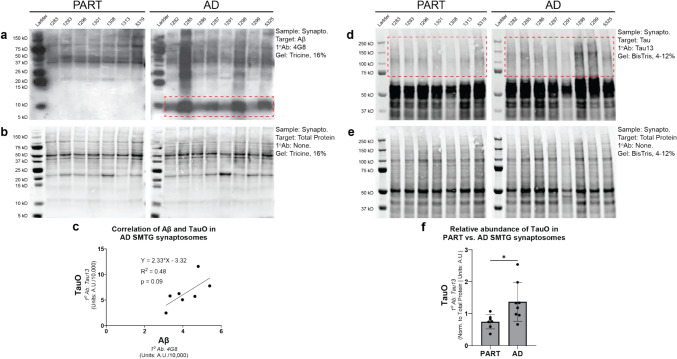
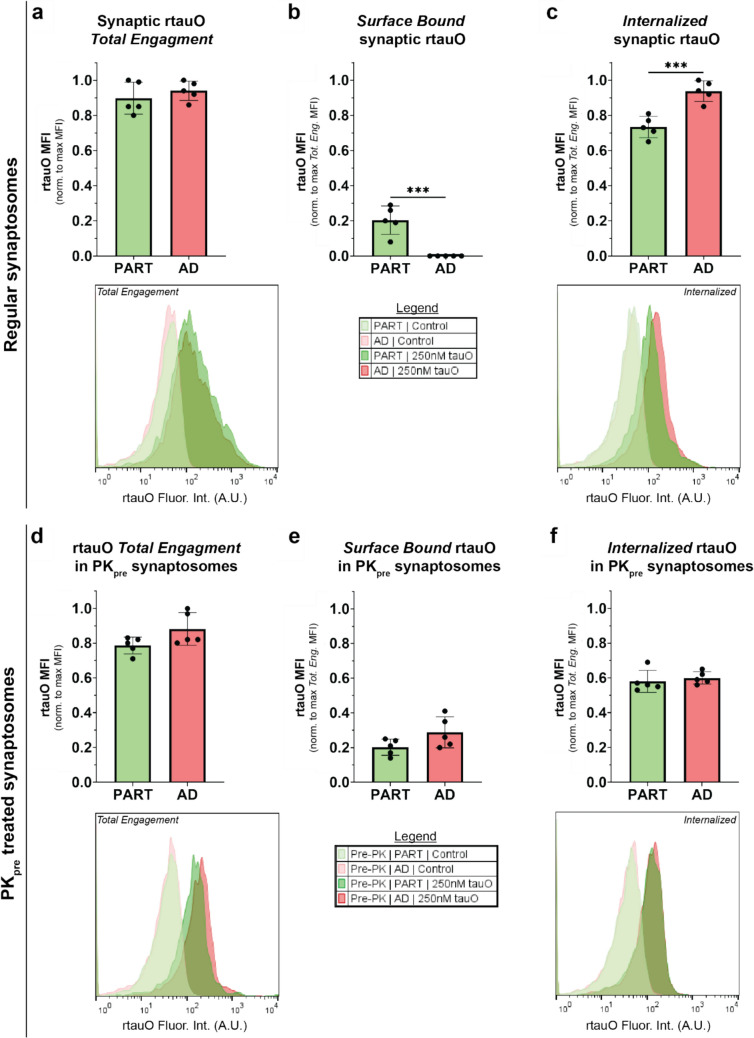
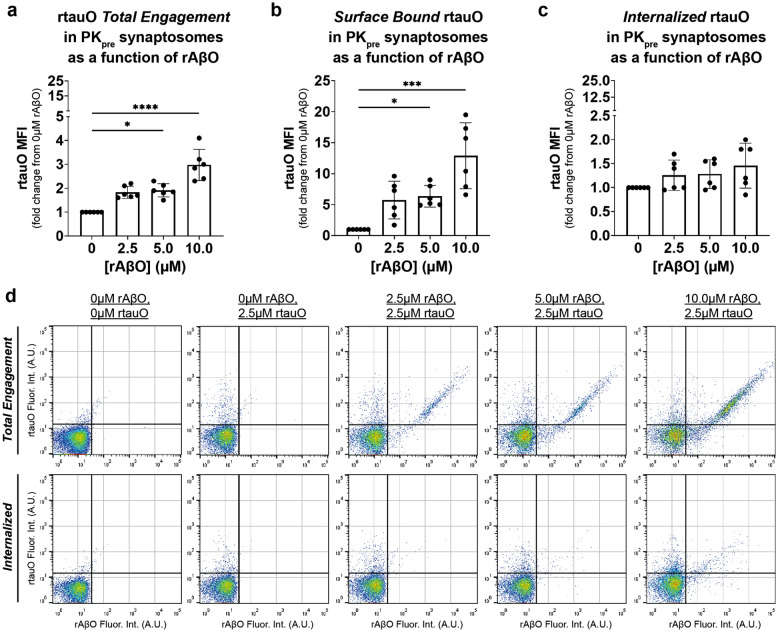
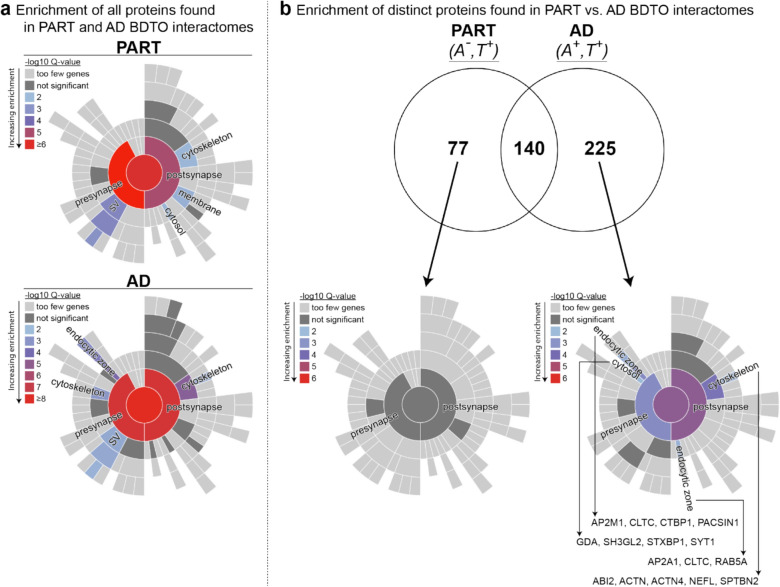
In Alzheimer's disease (AD), the propagation and spreading of CNS tau pathology closely correlates with cognitive decline, positioning tau as an attractive therapeutic target. Amyloid beta (Aβ) has been strongly implicated in driving tau spread, whereas primary tauopathies such as primary age-related tauopathy (PART)-which lack Aβ pathology-exhibit limited tau spread and minimal-to-no cognitive decline. Emerging evidence converges on a trans-synaptic mechanism of tau spread, facilitated by the transfer of misfolded tau aggregates (e.g. soluble oligomers). However, it is unclear whether Aβ oligomers modulate the binding and internalization of tau oligomers in human synapses. Our translationally focused paradigms utilize post-mortem brain specimens from Control, PART, and AD patients. Synaptosomes isolated from the temporal cortex of all three groups were incubated with preformed recombinant tauO (rtauO), ± preformed recombinant AβO (rAβO), and oligomer binding/internalization was quantified via flow cytometry following proteinase K (PK) digestion of surface-bound oligomers. TauO-synapse interactions were visualized using EM immunogold. Brain-derived tau oligomers (BDTO) from AD and PART PBS-soluble hippocampal fractions were co-immunoprecipitated and analyzed via mass spectrometry to compare synaptic tauO interactomes in primary and secondary tauopathies, thereby inferring the role of Aβ. AD synaptosomes, enriched in endogenous Aβ pathology, exhibited increased rtauO internalization compared to PART synaptosomes. This observation was mirrored in Control synaptosomes, where recombinant rAβO significantly increased rtauO binding and internalization. PK pre-treatment abolished this effect, implicating synaptic membrane proteins in AβO-mediated tauO internalization. While both PART and AD BDTO were broadly enriched in synaptic proteins, AD BDTO exhibited differential enrichment of endocytic proteins across pre- and post-synaptic compartments, whereas PART BDTO showed no significant synaptic enrichment. This study demonstrates that Aβ oligomers enhance tau oligomer binding and drive its internalization through synaptic membrane proteins. These findings offer novel mechanistic insights underlying pathological tau spreading directly within human synapses and emphasize the therapeutic potential of targeting Aβ-tau interactions.
Keywords: Aβ oligomers; Tau oligomers; Trans-synaptic tau spreading.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Conflict of interest: The authors declare that they have no financial or non-financial competing interests to disclose. Ethical approval and consent to participate: Postmortem frozen brain tissues used in this study were obtained through established material transfer agreements with the Alzheimer’s Disease Research Center (ADRC) at Sanders-Brown Center on Aging, University of Kentucky, and the Layton Aging and Alzheimer’s Disease Center (ADC) at Oregon Health and Science University. All donors were enrolled in ongoing brain aging studies, with informed consent obtained prior to participation, and all protocols adhered to Institutional Review Board (IRB) guidelines at each institution. All donor subject samples were de-identified before being provided to the University of Texas Medical Branch (UTMB); therefore, no approval was required from the UTMB IRB per CFR §46.101(a). Animals (mice) used procure EM immunogold images were housed in adherence with USDA standards and sacrificed in compliance with UTMB’s Institutional Animal Care and Use Committee (IACUC), as well as UTMB IRB, approved protocols.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
