

Internal feedback circuits among MEX-5, MEX-6, and PLK-1 maintain faithful patterning in the Caenorhabditis elegans embryo
- PMID: 39689170
- PMCID: PMC11670235
- DOI: 10.1073/pnas.2407517121
Internal feedback circuits among MEX-5, MEX-6, and PLK-1 maintain faithful patterning in the Caenorhabditis elegans embryo
Abstract
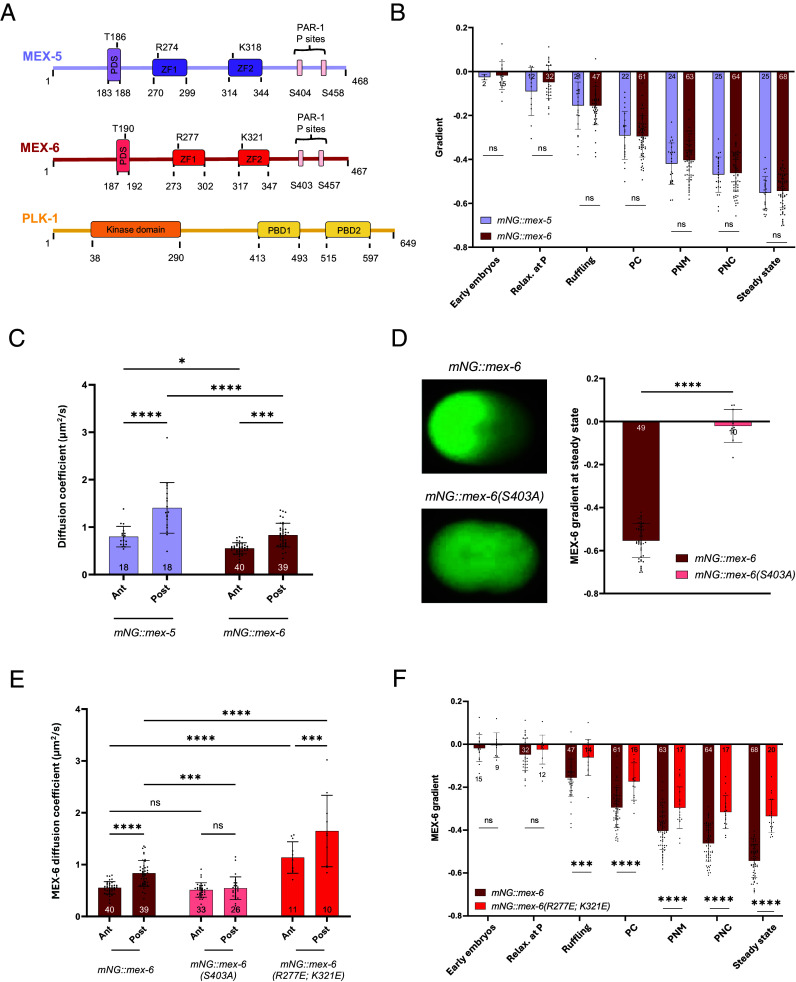
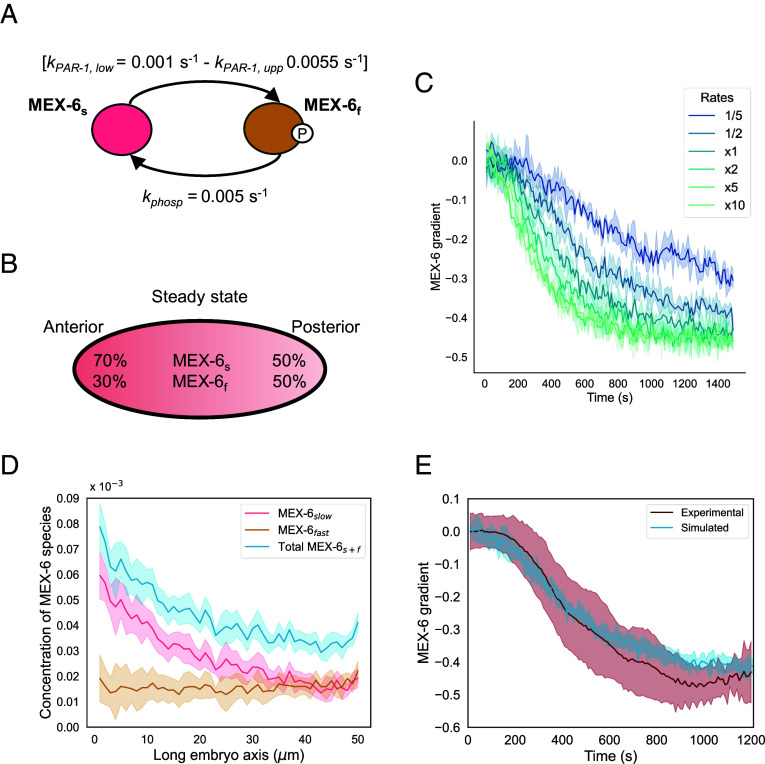
Proteins become asymmetrically distributed in the one-cell Caenorhabditis elegans embryo thanks to reaction-diffusion mechanisms that are often entangled in complex feedback loops. Cortical polarity drives the enrichment of the RNA-binding proteins MEX-5 and MEX-6 in the anterior cytoplasm through concentration gradients. MEX-5 and MEX-6 promote the patterning of other cytoplasmic factors, including that of the anteriorly enriched mitotic polo-like kinase PLK-1, but also contribute to proper cortical polarity. The gradient of MEX-5 forms through a differential-diffusion mechanism. How MEX-6 establishes a gradient and how MEX-5 and MEX-6 regulate cortical polarity is not known. Here, we reveal that the two MEX proteins develop concentration asymmetries via similar mechanisms, but despite their strong sequence homology, they differ in terms of how their concentration gradients are regulated. We find that PLK-1 promotes the enrichment of MEX-5 and MEX-6 at the anterior through different circuits: PLK-1 influences the MEX-5 gradient indirectly by regulating cortical polarity while it modulates the formation of the gradient of MEX-6 through its physical interaction with the protein. We thus propose a model in which PLK-1 mediates protein circuitries between MEX-5, MEX-6, and cortical proteins to faithfully establish and maintain polarity.
Keywords: MEX-5, MEX-6, PLK-1; Monte Carlo simulations; RNA binding proteins; polarity feedback loops; reaction–diffusion.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures



References
-
- Brown G. C., Kholodenko B. N., Spatial gradients of cellular phospho-proteins. FEBS Lett. 457, 452–454 (1999). - PubMed
-
- Cheeks R. J., et al. , C. elegans PAR proteins function by mobilizing and stabilizing asymmetrically localized protein complexes. Curr. Biol. 14, 851–862 (2004). - PubMed
-
- Munro E., Nance J., Priess J. R., Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior–posterior polarity in the early C. elegans embryo. Dev. Cell 7, 413–424 (2004). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
