

Molecular mechanism targeting condensin for chromosome condensation
- PMID: 39690240
- PMCID: PMC11791182
- DOI: 10.1038/s44318-024-00336-6
Molecular mechanism targeting condensin for chromosome condensation
Abstract
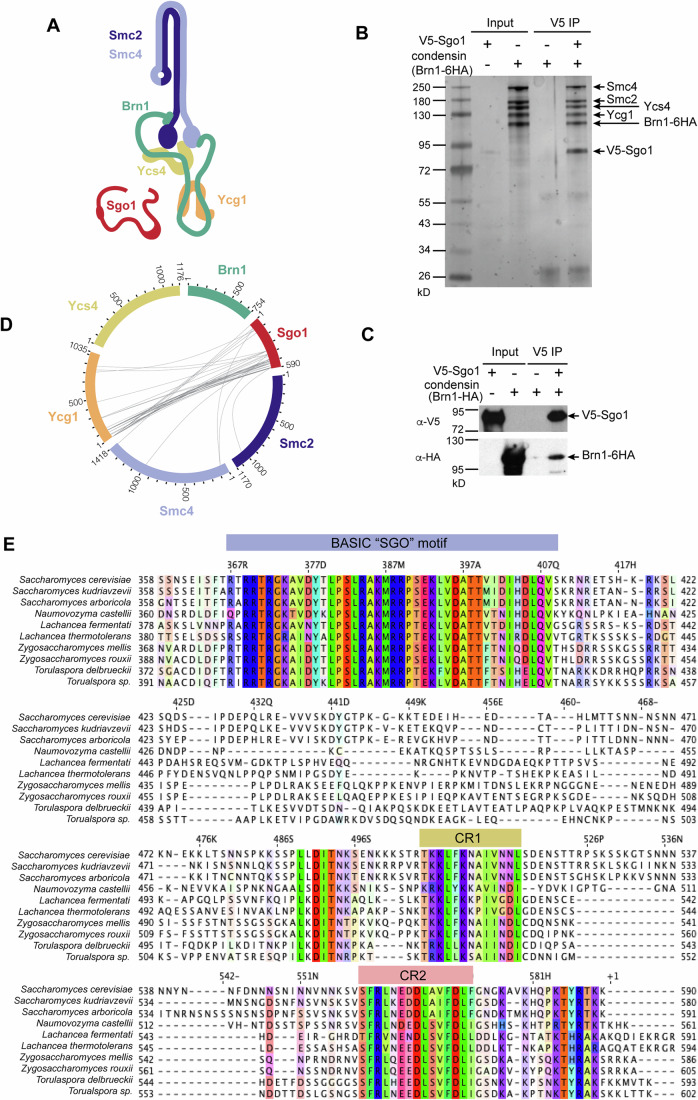
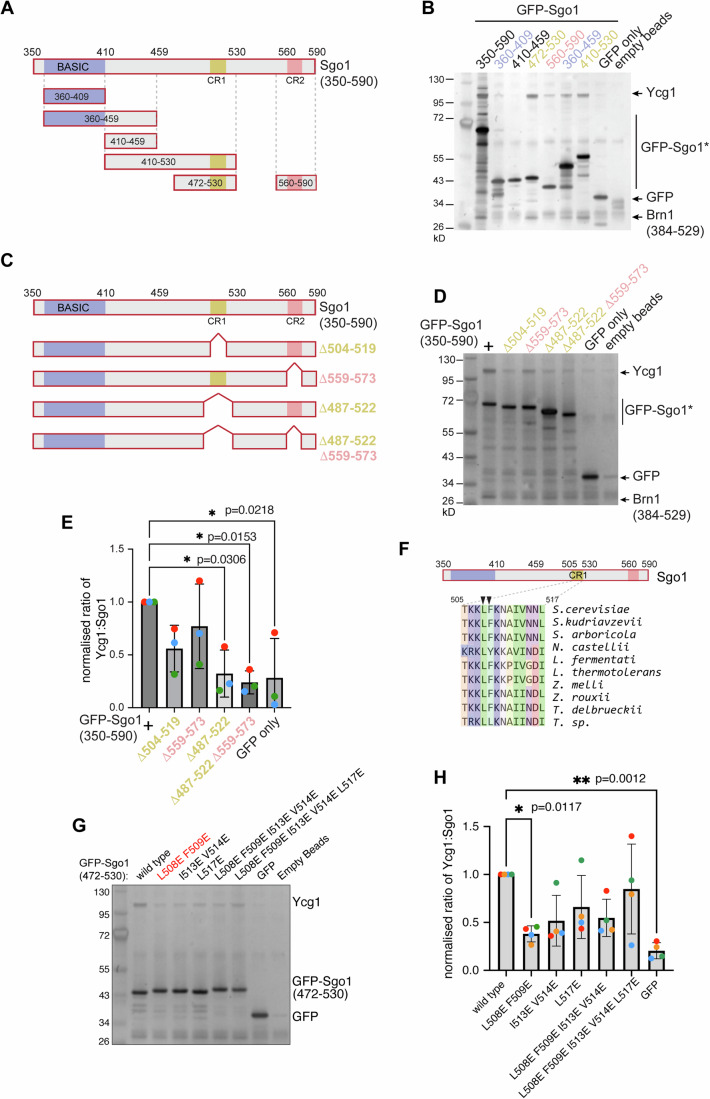
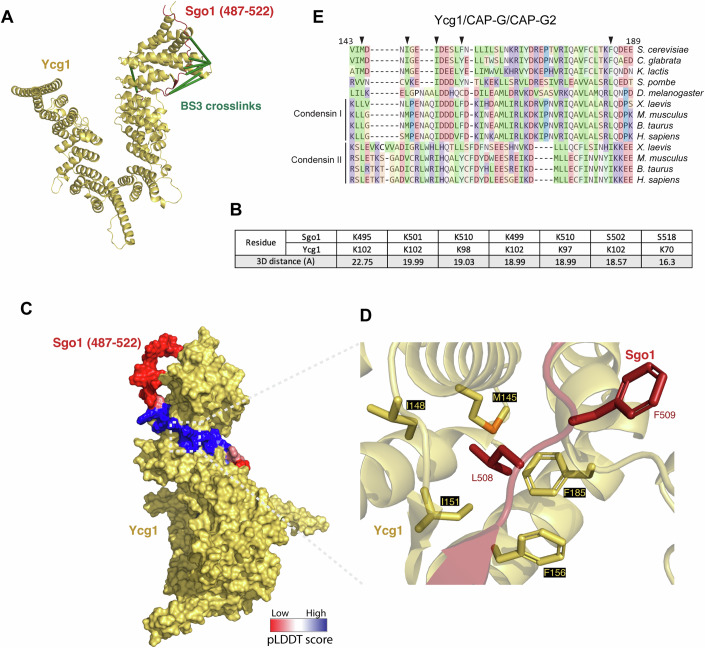
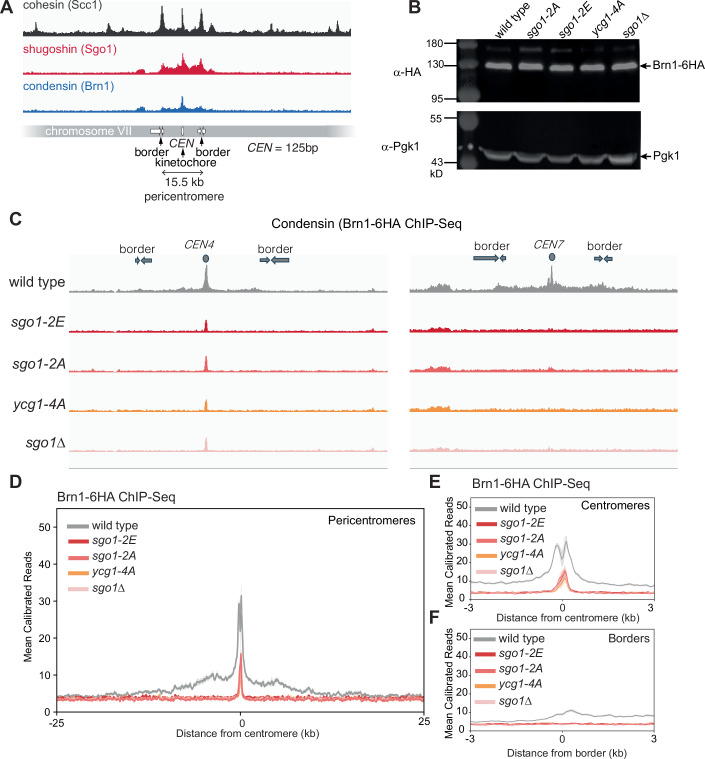
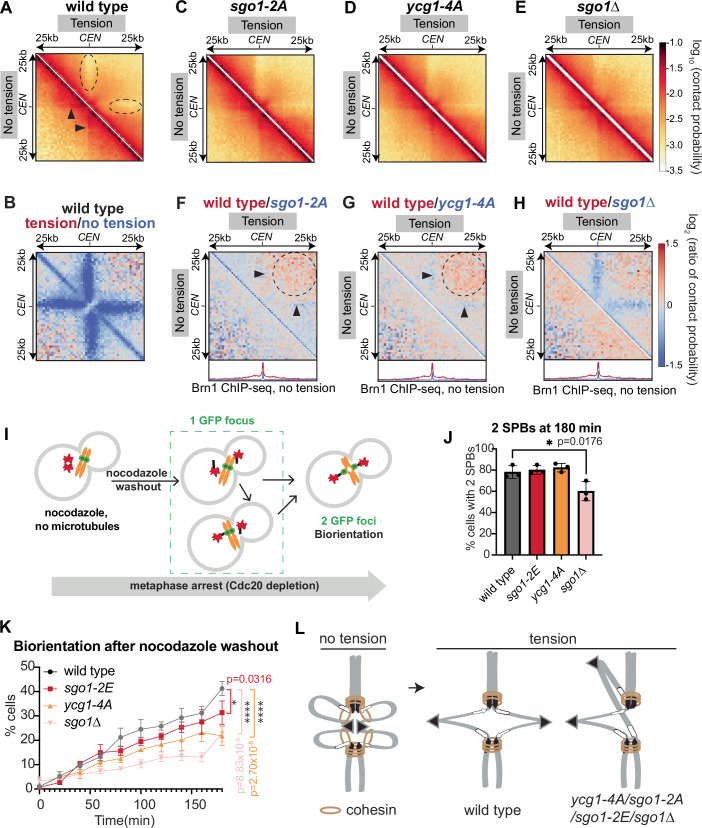
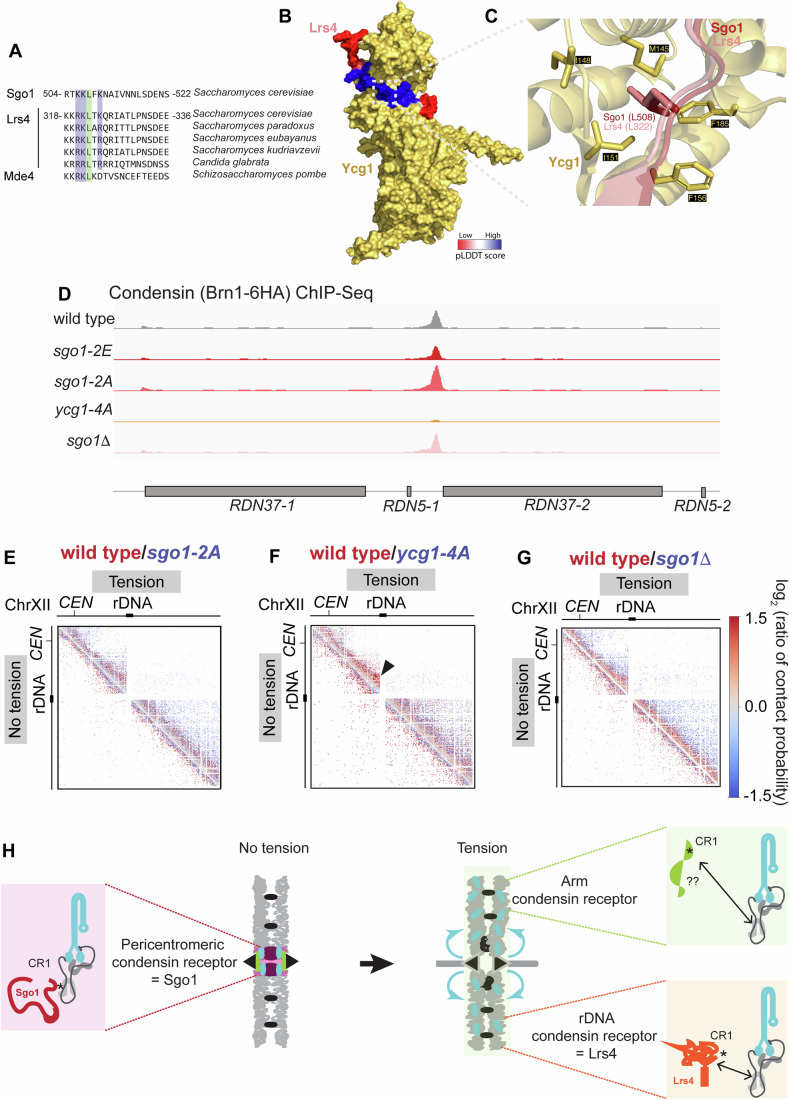
Genomes are organised into DNA loops by the Structural Maintenance of Chromosomes (SMC) proteins. SMCs establish functional chromosomal sub-domains for DNA repair, gene expression and chromosome segregation, but how SMC activity is specifically targeted is unclear. Here, we define the molecular mechanism targeting the condensin SMC complex to specific chromosomal regions in budding yeast. A conserved pocket on the condensin HAWK subunit Ycg1 binds to chromosomal receptors carrying a related motif, CR1. In early mitosis, CR1 motifs in receptors Sgo1 and Lrs4 recruit condensin to pericentromeres and rDNA, to facilitate sister kinetochore biorientation and rDNA condensation, respectively. We additionally find that chromosome arm condensation begins as sister kinetochores come under tension, in a manner dependent on the Ycg1 pocket. We propose that multiple CR1-containing proteins recruit condensin to chromosomes and identify several additional candidates based on their sequence. Overall, we uncover the molecular mechanism that targets condensin to functionalise chromosomal domains to achieve accurate chromosome segregation during mitosis.
Keywords: Condensin; Lrs4; Pericentromeres; Shugoshin; rDNA.
© 2024. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. The authors declare no competing interests.
Figures







References
-
- Baxter J, Sen N, Martínez VL, De Carandini MEM, Schvartzman JB, Diffley JFX, Aragon L et al (2011) Positive supercoiling of mitotic DNA drives decatenation by topoisomerase II in eukaryotes. Science 331:1328–1332 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
