

GABAAR-PPT1 palmitoylation homeostasis controls synaptic transmission and circuitry oscillation
- PMID: 39695089
- PMCID: PMC11655527
- DOI: 10.1038/s41398-024-03206-1
GABAAR-PPT1 palmitoylation homeostasis controls synaptic transmission and circuitry oscillation
Abstract
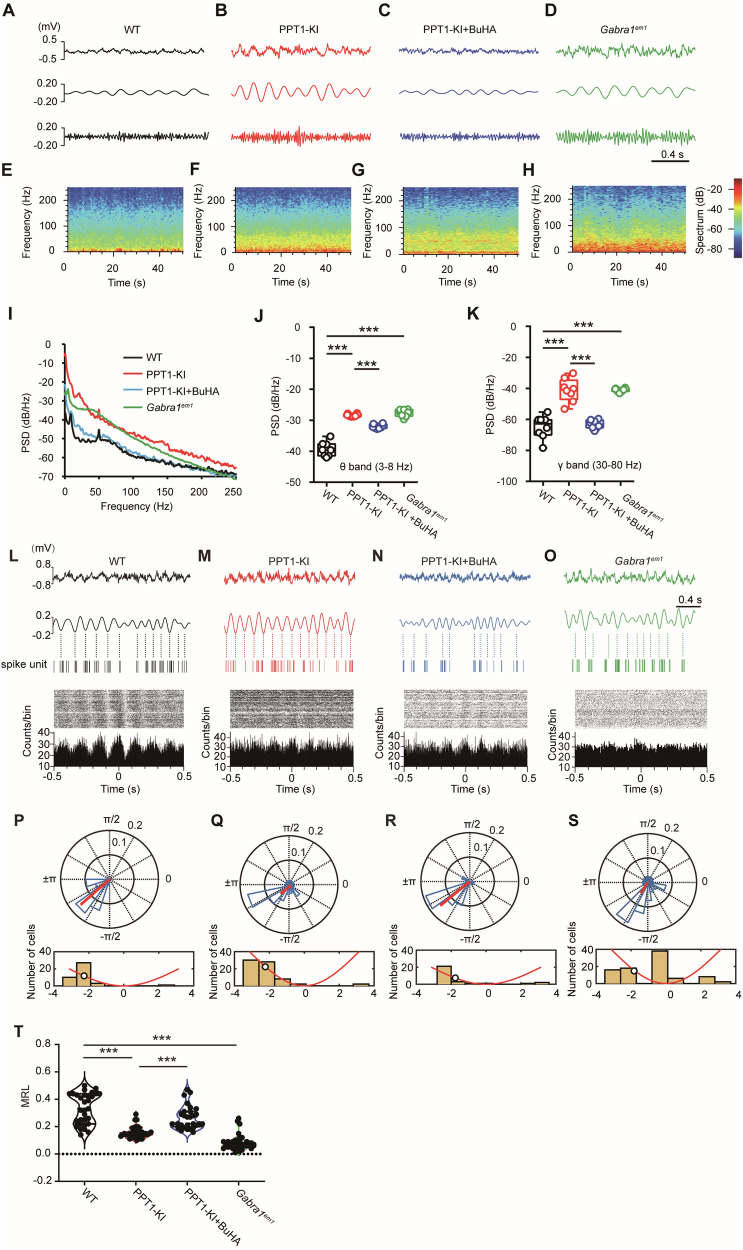
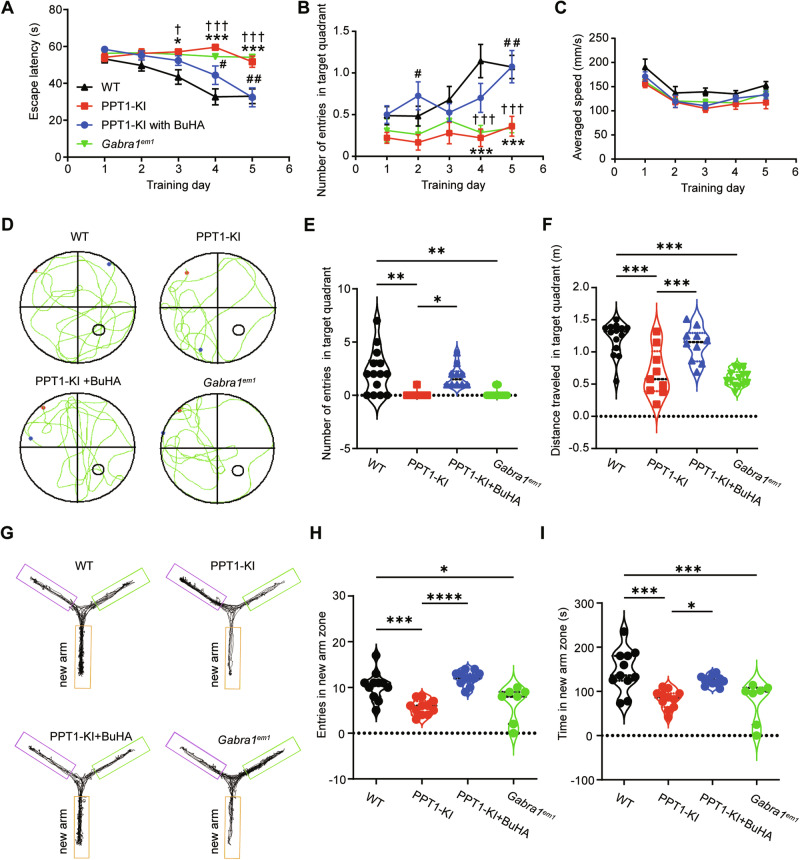
The infantile neuronal ceroid lipofuscinosis, also called CLN1 disease, is a fatal neurodegenerative disease caused by mutations in the CLN1 gene encoding palmitoyl protein thioesterase 1 (PPT1). Identifying the depalmitoylation substrates of PPT1 is crucial for understanding CLN1 disease. In this study, we found that GABAAR, the critical synaptic protein essential for inhibitory neurotransmission, is a substrate of PPT1. PPT1 depalmitoylates GABAAR α1 subunit at Cystein-260, while binding to Cystein-165 and -179. Mutations of PPT1 or its GABAAR α1 subunit binding site enhanced inhibitory synaptic transmission and strengthened oscillations powers but disrupted phase coupling in CA1 region and impaired learning and memory in 1- to 2-months-old PPT1-deficient and Gabra1em1 mice. Our study highlights the critical role of PPT1 in maintaining GABAAR palmitoylation homeostasis and reveals a previously unknown molecular pathway in CLN1 diseases induced by PPT1 mutations.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval and consent to participate: The experiments did not involve tissues from humans. Animal use and procedures were performed according to the regulations and requirements of XXMU Animal Ethics Committee (No. XYLL2021053).
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
