

Unweaving the Cognitive Map: A Personal History
- PMID: 39698925
- PMCID: PMC11656510
- DOI: 10.1002/hipo.23674
Unweaving the Cognitive Map: A Personal History
Abstract
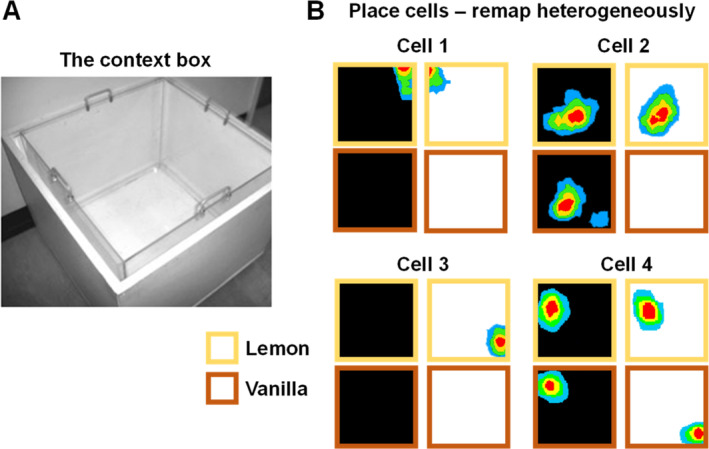
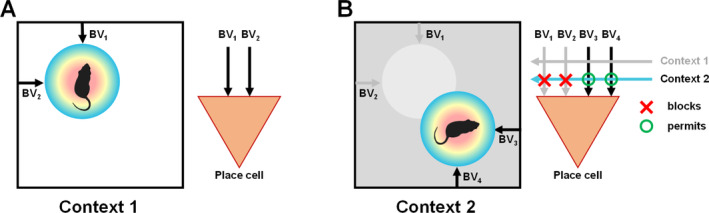
I have been incredibly fortunate to have worked in the field of hippocampal spatial coding during three of its most exciting decades, the 1990s, 2000s, and 2010s. During this time I had a ringside view of some of the foundational discoveries that were made which have transformed our understanding of the hippocampal system and its role in cognition (especially spatial cognition) and memory. These discoveries inspired me in my own lab over the years to pursue three broad lines of enquiry-3D spatial encoding, context and the sense of direction-which are outlined here. If some of my personal recollections are a little inaccurate (such is the nature of episodic memory!) I apologize in advance.
Keywords: configural learning; context; head direction cells; hippocampus; place cells; spatial cognition.
© 2024 The Author(s). Hippocampus published by Wiley Periodicals LLC.
Conflict of interest statement
The author declares the following competing interests: K.J.J. is a non‐shareholding director of Axona Ltd., a company that makes and sells tetrode‐based electrophysiology products.
Figures







Similar articles
-
From British Associationism to the Hippocampal Cognitive Map: A Personal View From a Ringside Seat at the Cognitive/PDP Revolution.Hippocampus. 2025 Jan;35(1):e23662. doi: 10.1002/hipo.23662. Hippocampus. 2025. PMID: 39665512 Free PMC article. Review.
-
Memory and Space: Towards an Understanding of the Cognitive Map.J Neurosci. 2015 Oct 14;35(41):13904-11. doi: 10.1523/JNEUROSCI.2618-15.2015. J Neurosci. 2015. PMID: 26468191 Free PMC article.
-
Time, memory, and the legacy of Howard Eichenbaum.Hippocampus. 2019 Mar;29(3):146-161. doi: 10.1002/hipo.23007. Epub 2018 Sep 11. Hippocampus. 2019. PMID: 29979481 Review.
-
Explicit Memory, Implicit Memory, and the Hippocampus: Insights From Early Neuroimaging Studies.Hippocampus. 2025 Jan;35(1):e23657. doi: 10.1002/hipo.23657. Hippocampus. 2025. PMID: 39644280
-
Episodic Aspects of a Path Navigated Through Hippocampal Neurobiology.Hippocampus. 2025 Jan;35(1):e23672. doi: 10.1002/hipo.23672. Hippocampus. 2025. PMID: 39719302 Review.
Cited by
-
Towards an Understanding of the Dentate Gyrus Hilus.Hippocampus. 2025 Jan;35(1):e23677. doi: 10.1002/hipo.23677. Hippocampus. 2025. PMID: 39721944 Review.
References
-
- Anderson, M. I. , Killing S., Morris C., et al. 2006. “Behavioral Correlates of the Distributed Coding of Spatial Context.” Hippocampus 16, no. 9: 730–742. - PubMed
-
- Barry, C. , Hayman R., Burgess N., and Jeffery K. J.. 2007. “Experience‐Dependent Rescaling of Entorhinal Grids.” Nature Neuroscience 10, no. 6: 682–684. - PubMed
-
- Bostock, E. , Muller R. U., and Kubie J. L.. 1991. “Experience‐Dependent Modifications of Hippocampal Place Cell Firing.” Hippocampus 1, no. 2: 193–205. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
