

Pharmacological blocking of spinal GABAA receptors in monkeys reduces sensory transmission to the spinal cord, thalamus, and cortex
- PMID: 39700009
- PMCID: PMC12442854
- DOI: 10.1016/j.celrep.2024.115100
Pharmacological blocking of spinal GABAA receptors in monkeys reduces sensory transmission to the spinal cord, thalamus, and cortex
Abstract
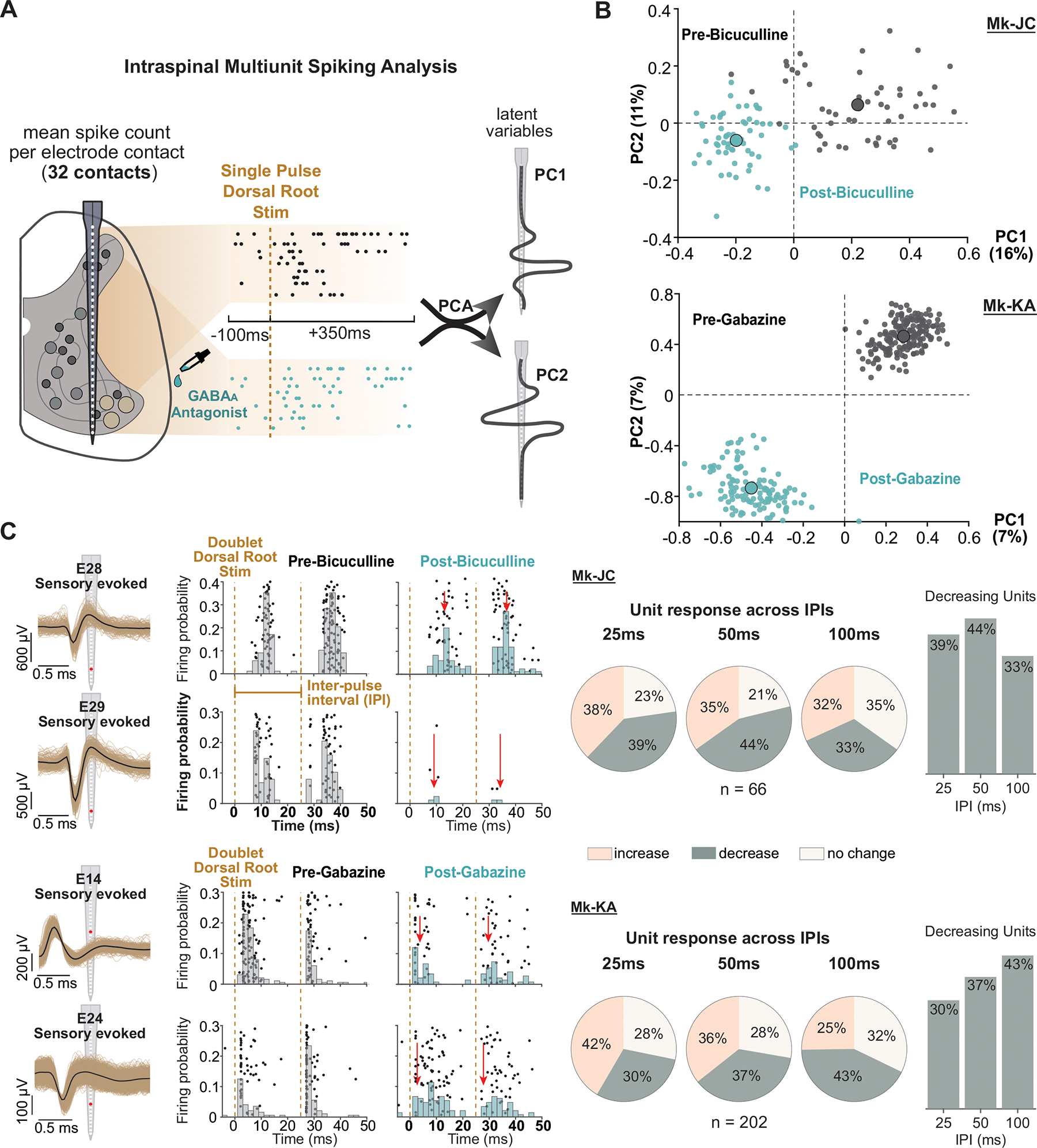
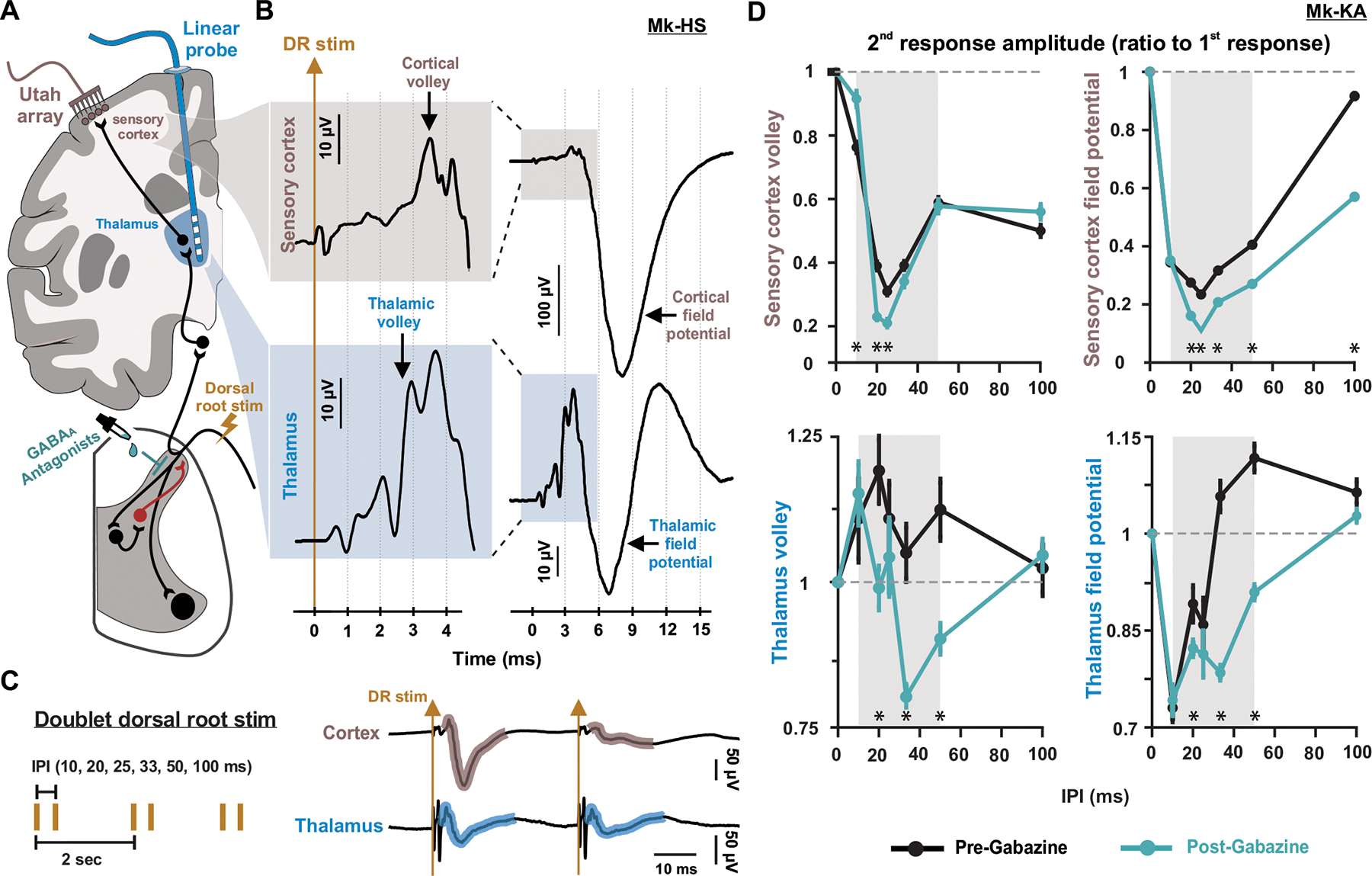
A century of research established that GABA inhibits proprioceptive inputs presynaptically to sculpt spinal neural inputs into skilled motor output. Recent results in mice challenged this theory by showing that GABA can also facilitate action potential conduction in proprioceptive afferents. Here, we tackle this controversy in monkeys, the most human-relevant animal model, and show that GABAA receptors (GABAARs) indeed facilitate sensory inputs to spinal motoneurons and interneurons and that this mechanism also influences sensory transmission to supraspinal centers. We performed causal manipulations of GABAARs with intrathecal pharmacology in anesthetized monkeys while recording electrical signals in the muscles, spinal cord, thalamus, and cortex. We show that blocking GABAARs suppresses spinal reflexes to hand muscles, sensory-evoked single-unit firing in the spinal cord, and sensory-evoked potentials in the thalamus and somatosensory cortex. Our results portray a sophisticated picture of presynaptic modulation of sensory inputs by GABA in the spinal cord.
Keywords: CP: Cell biology; CP: Neuroscience; GABA; GABA(A) receptors; hand; monkeys; presynaptic inhibition; primary afferent depolarization; sensory transmission; somatosensory cortex; spinal reflexes; thalamus.
Copyright © 2024. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


References
-
- Gotch F, and Horsley VAH (1891). VI. Croonian Lecture.—On the mammalian nervous system, its functions, and their localisation determined by an electrical method. Philosophical Transactions of the Royal Society of London. , 267–526. 10.1098/rstb.1891.0006. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
