

NPR1 promotes blue light-induced plant photomorphogenesis by ubiquitinating and degrading PIF4
- PMID: 39700134
- PMCID: PMC11670118
- DOI: 10.1073/pnas.2412755121
NPR1 promotes blue light-induced plant photomorphogenesis by ubiquitinating and degrading PIF4
Abstract
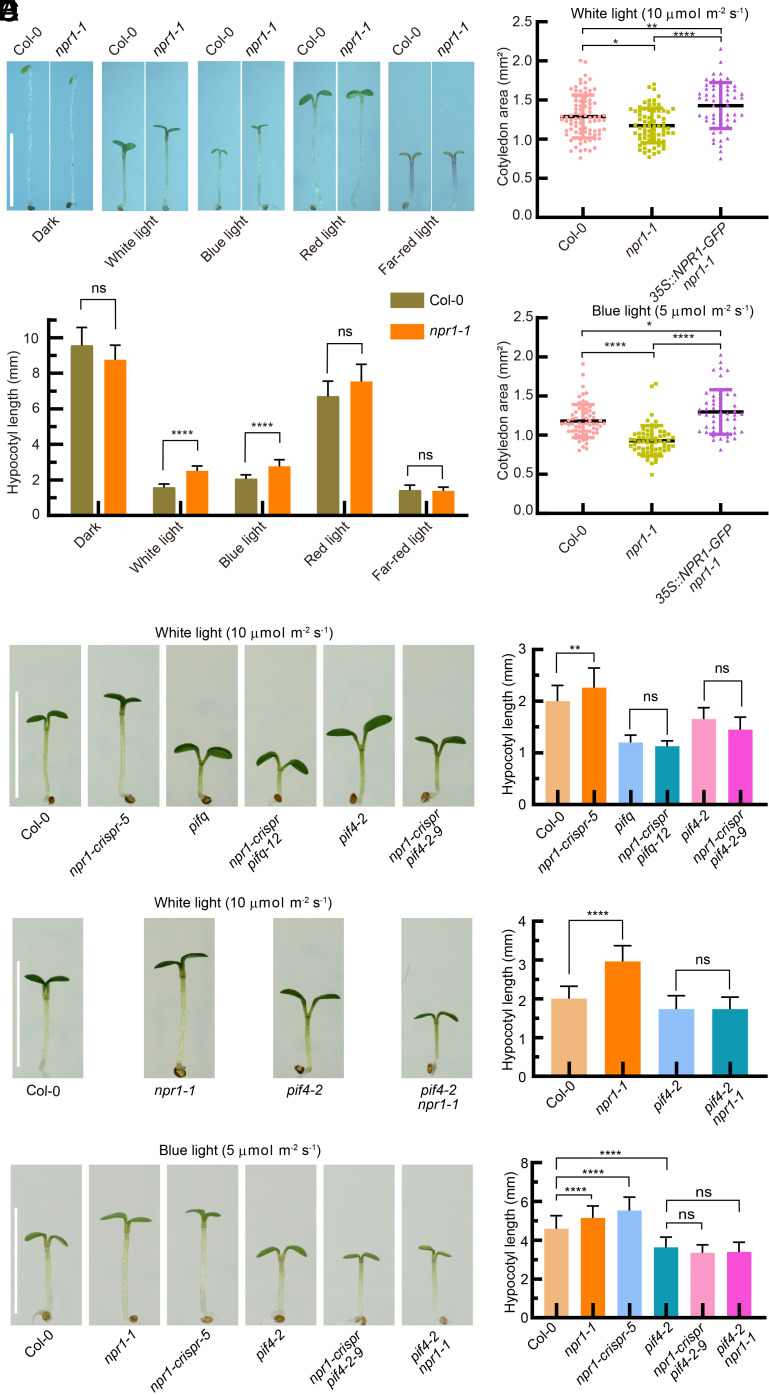
Light is a major determinant of plant growth and survival. NONEXPRESSER OF PATHOGENESIS-RELATED GENES 1 (NPR1) acts as a receptor for salicylic acid (SA) and serves as the key regulator of SA-mediated immune responses. However, the mechanisms by which plants integrate light and SA signals in response to environmental changes, as well as the role of NPR1 in regulating plant photomorphogenesis, remain poorly understood. This study shows that SA promotes plant photomorphogenesis by regulating PHYTOCHROME INTERACTING FACTOR 4 (PIF4). Specifically, NPR1 promotes photomorphogenesis under blue light by facilitating the degradation of PIF4 through light-induced polyubiquitination. NPR1 acts as a substrate adaptor for the CULLIN3-based E3 ligase, which ubiquitinates PIF4 at Lys129, Lys252, and Lys428, and leading to PIF4 degradation via the 26S proteasome pathway. Genetically, PIF4 is epistatic to NPR1 in the regulation of blue light-induced photomorphogenesis, suggesting it acts downstream of NPR1. Furthermore, cryptochromes mediate the polyubiquitination of PIF4 by NPR1 in response to blue light by promoting the interaction and ubiquitination between NPR1 and PIF4. Transcriptome analysis revealed that under blue light, NPR1 and PIF4 coordinately regulate numerous downstream genes related to light and auxin signaling pathways. Overall, these findings unveil a role for NPR1 in photomorphogenesis, highlighting a mechanism for posttranslational regulation of PIF4 in response to blue light. This mechanism plays a pivotal role in the fine-tuning of plant development, enabling plants to adapt to complex environmental changes.
Keywords: NPR1; PIF4; photomorphogenesis; salicylic acid; ubiquitination.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures





References
-
- Deng X. W., et al. , COP1, an Arabidopsis regulatory gene, encodes a protein with both a zinc-binding motif and a G beta homologous domain. Cell 71, 791–801 (1992). - PubMed
-
- Josse E. M., Halliday K. J., Skotomorphogenesis: The dark side of light signalling. Curr. Biol. 18, R1144–R1146 (2008). - PubMed
MeSH terms
Substances
Grants and funding
- 5232010/| Natural Science Foundation of Beijing Municipality (Beijing Natural Science Foundation)
- 32170292/MOST | National Natural Science Foundation of China (NSFC)
- PC2023B02010/Pinduoduo-China Agricultural University Research Fund
- 2022RC042/MOE | Chinese Universities Scientific Fund
- 2021QNRC001/Young Elite Scientists Sponsorship Program by China Association of Science & Technology
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
