

Coupling antigens from multiple subtypes of influenza can broaden antibody and T cell responses
- PMID: 39700292
- PMCID: PMC12036609
- DOI: 10.1126/science.adi2396
Coupling antigens from multiple subtypes of influenza can broaden antibody and T cell responses
Abstract
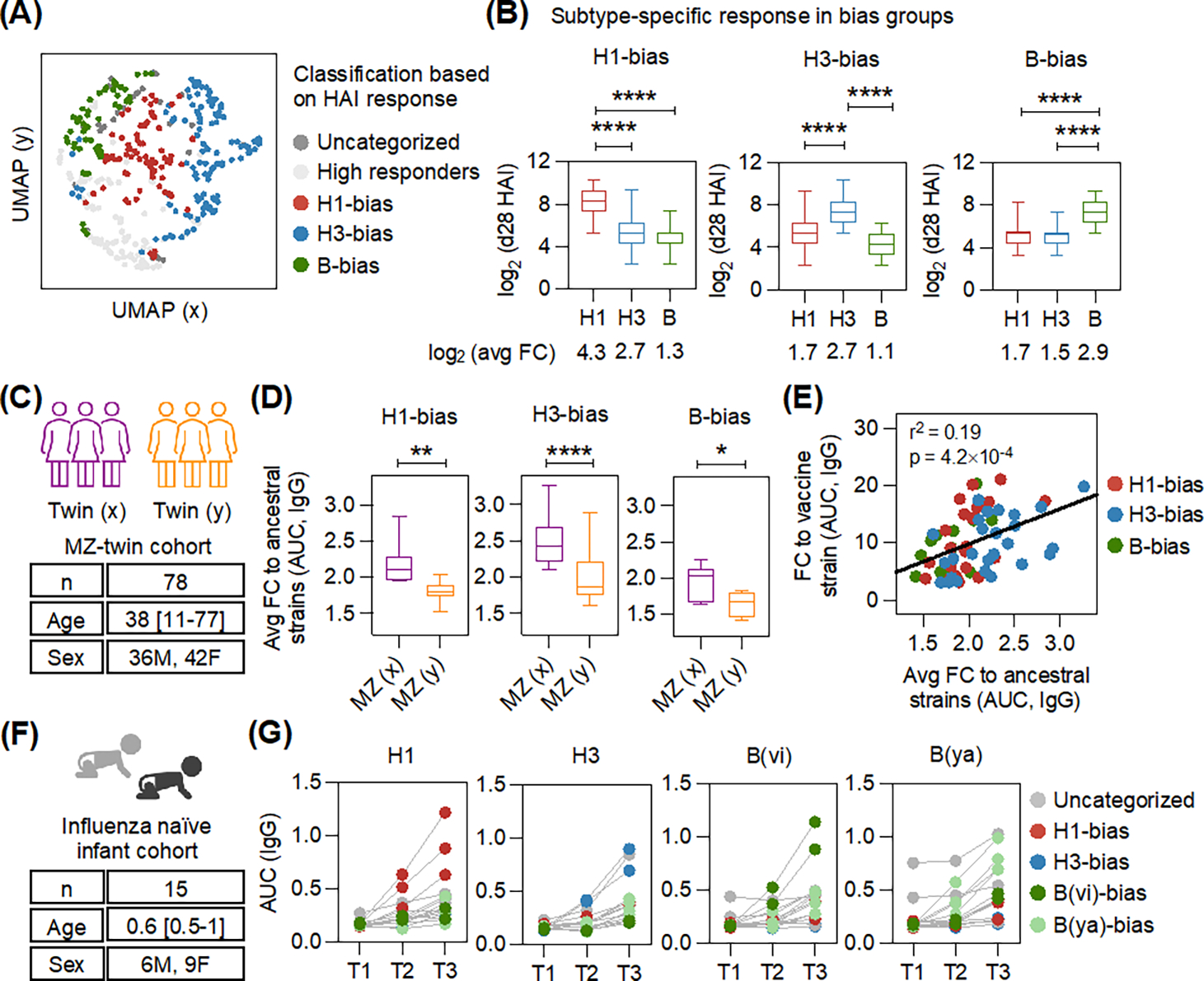
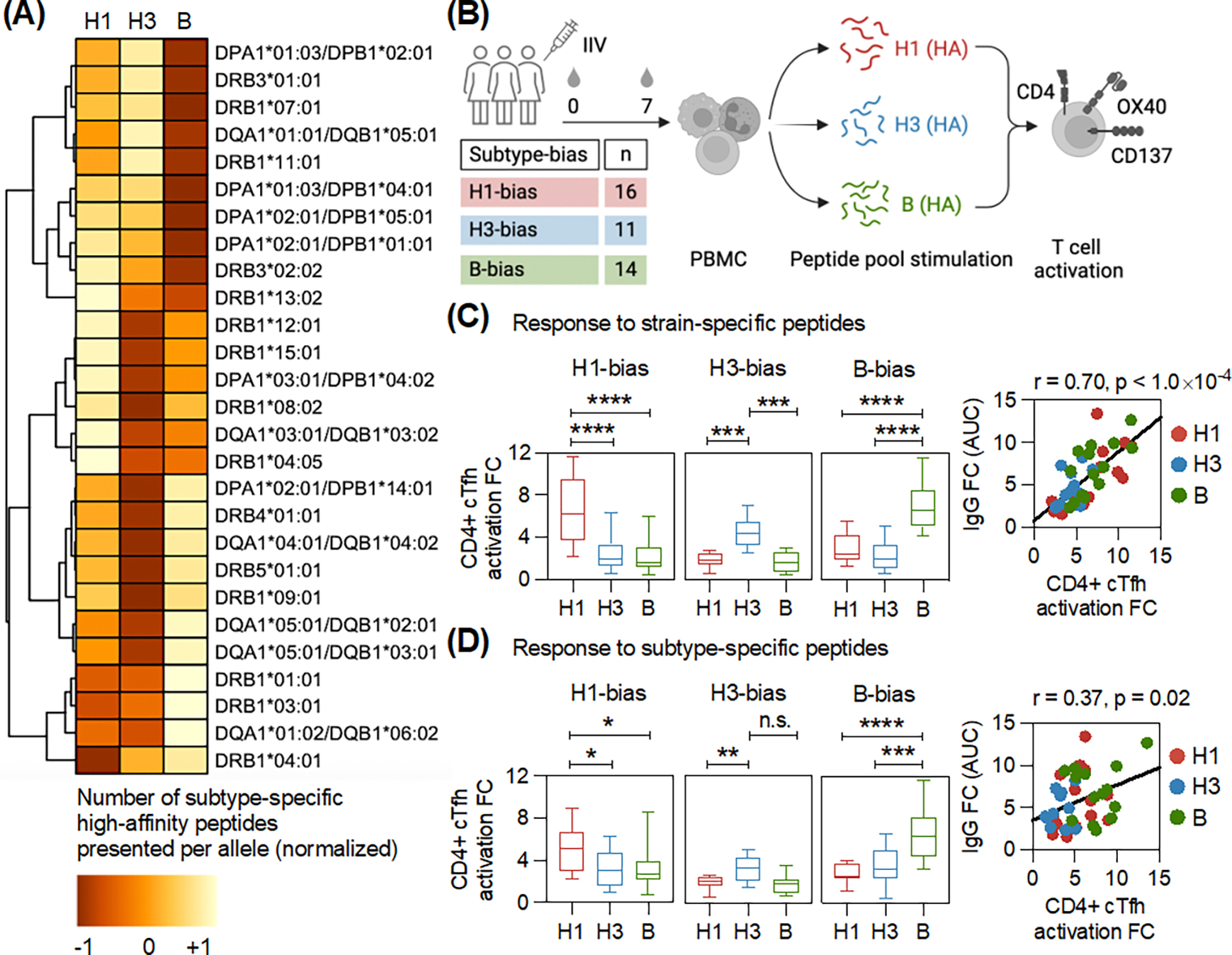
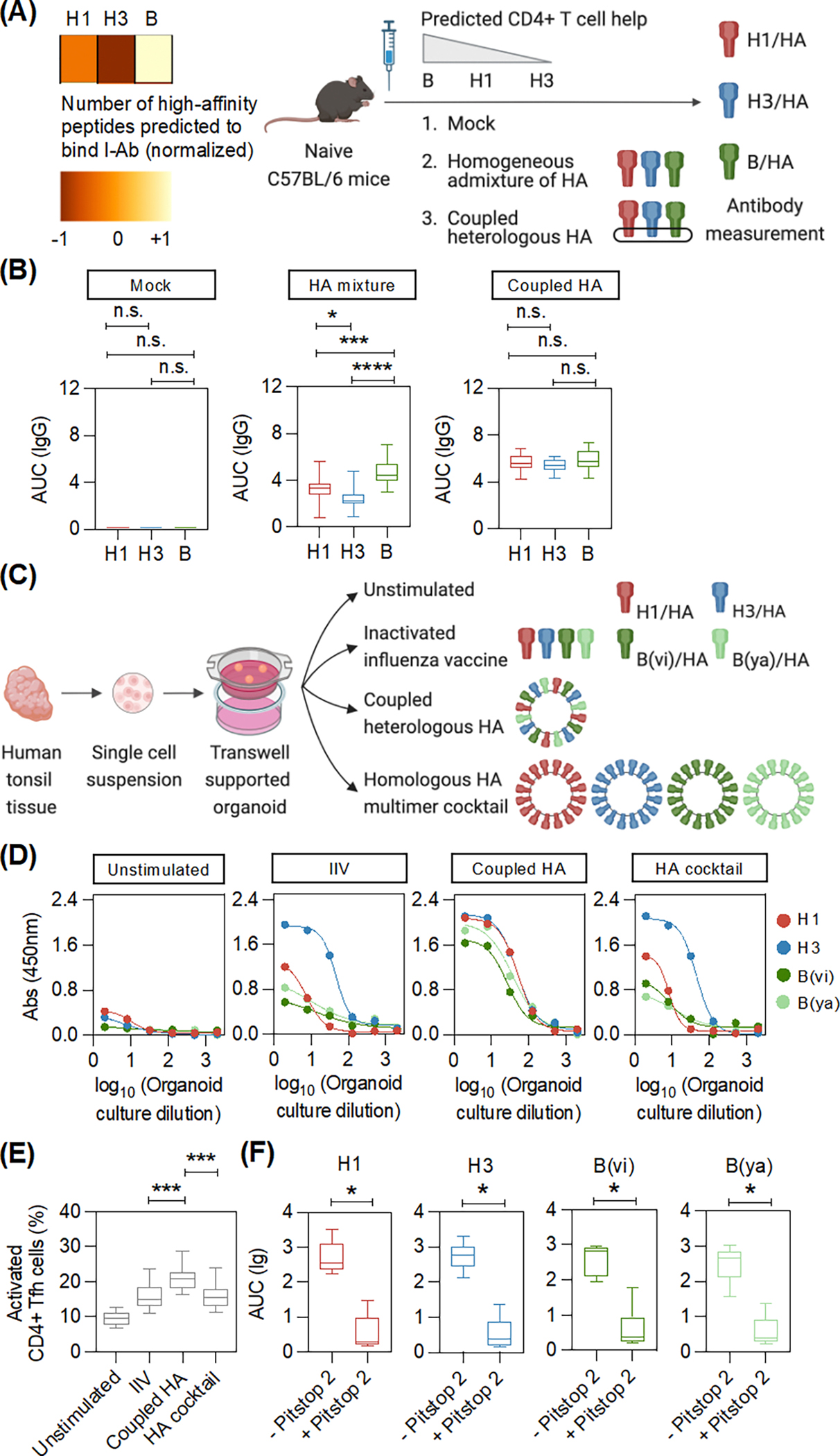
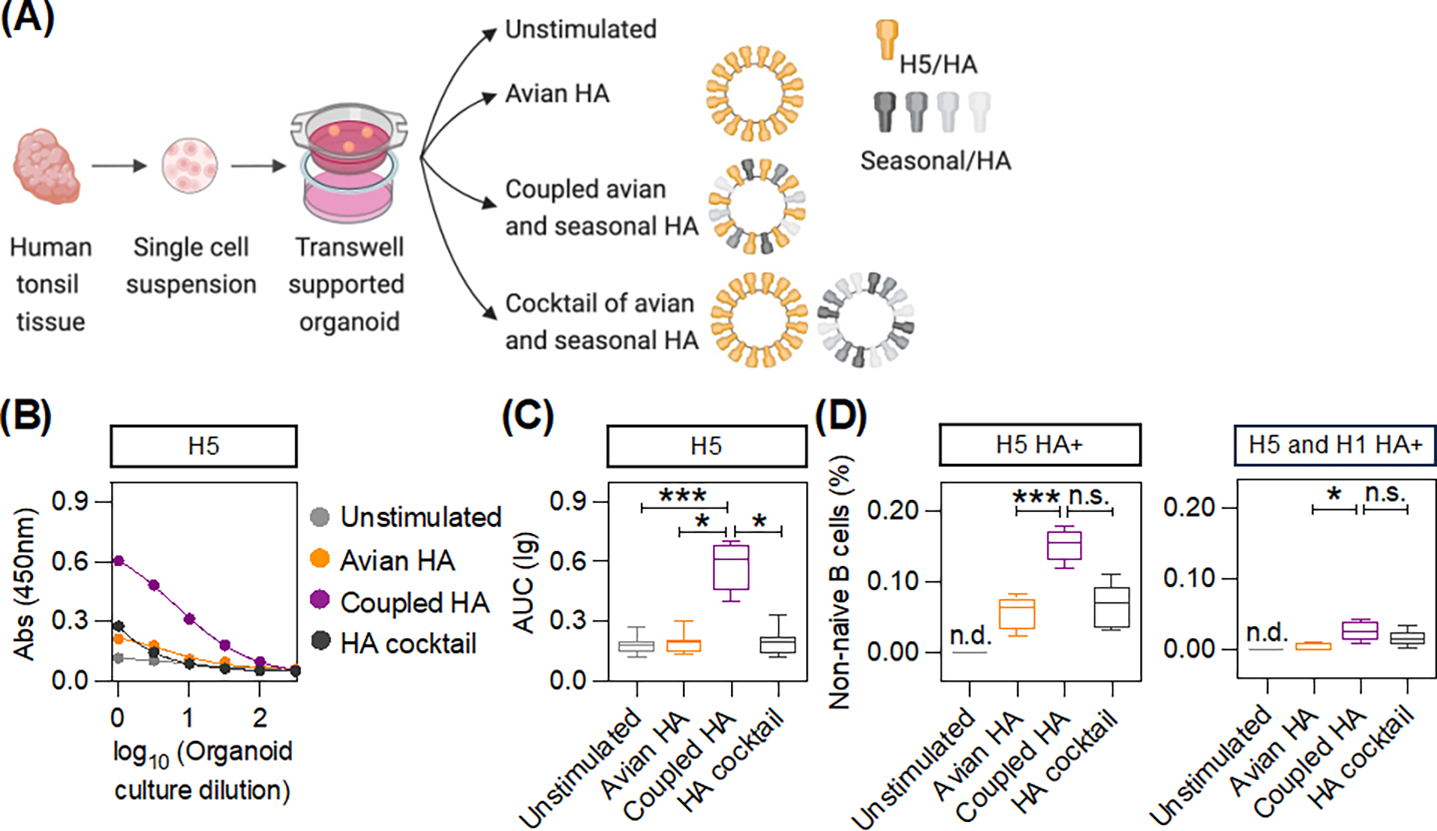
The seasonal influenza vaccine contains strains of viruses from distinct subtypes that are grown independently and then combined. However, most individuals exhibit a more robust response to one of these strains and thus are vulnerable to infection by others. By studying a monozygotic twin cohort, we found that although prior exposure is a factor, host genetics are a stronger driver of subtype bias to influenza viral strains. We found that covalent coupling of heterologous hemagglutinin (HA) from different viral strains could largely eliminate subtype bias in an animal model and in a human tonsil organoid system. We proposed that coupling of heterologous antigens improves antibody responses across influenza strains by broadening T cell help, and we found that using this approach substantially improved the antibody response to avian influenza HA.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
