

Cryo-EM structures of the full-length human contactin-2
- PMID: 39702996
- PMCID: PMC11796320
- DOI: 10.1111/febs.17364
Cryo-EM structures of the full-length human contactin-2
Abstract
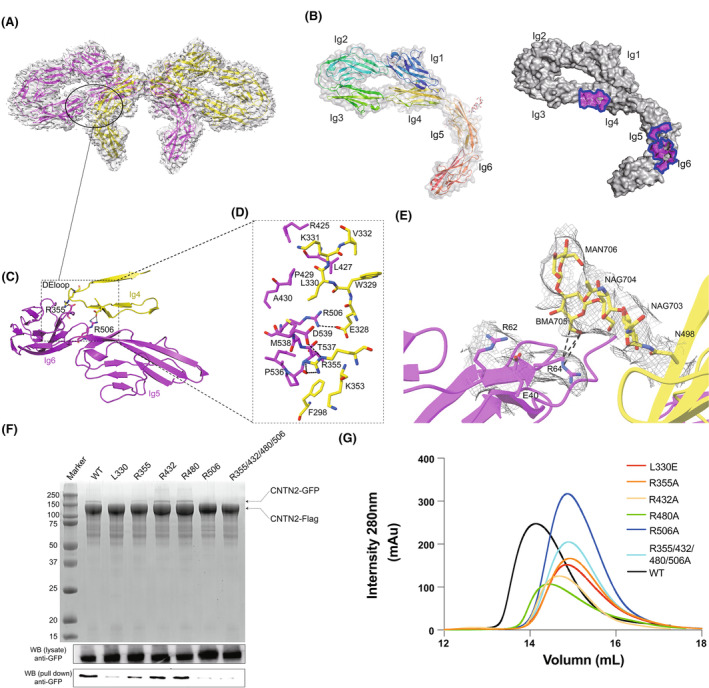
Contactin-2 (CNTN2), an immunoglobulin cell adhesion molecule (IgCAM) expressed on the neural cell surface, regulates the formation of myelin sheaths, facilitates communication between neurons and axoglial cells, and coordinates the migration of neural cells. However, the assembly of full-length CNTN2 is still not fully elucidated. Here, we found that the full-length human CNTN2 forms a concentration-dependent homodimer. We further determined the cryo-EM structures of the full-length CNTN2, revealing a novel bowknot-shaped scaffold constituted of the Ig1-6 repeats from two protomers, with the flexible ribbon-like FNIII repeats extending outward in opposite directions. The Ig1-6 domains, rather than the previously proposed Ig1-4 domains, have an indispensable role in mediating CNTN2-dependent cell adhesion and clustering. Moreover, structure-guided mutagenesis analyses supported the idea that CNTN2 homodimerization observed in our structure is essential for cell adhesion. Our findings offer novel insights into the mechanism through which CNTN2 forms a homodimer to maintain cell-cell contacts in the nervous system.
Keywords: cell adhesion; contactin‐2; cryo‐EM; dimerization; full‐length.
© 2024 The Author(s). The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors affirm that the research was conducted without any commercial or financial relationships that might be perceived as a potential conflict of interest.
Figures






Similar articles
-
Molecular mechanism of contactin 2 homophilic interaction.Structure. 2024 Oct 3;32(10):1652-1666.e8. doi: 10.1016/j.str.2024.06.004. Epub 2024 Jul 4. Structure. 2024. PMID: 38968938
-
New insights into the roles of the contactin cell adhesion molecules in neural development.Adv Neurobiol. 2014;8:165-94. doi: 10.1007/978-1-4614-8090-7_8. Adv Neurobiol. 2014. PMID: 25300137 Review.
-
The role of Gpi-anchored axonal glycoproteins in neural development and neurological disorders.Mol Cell Neurosci. 2017 Jun;81:49-63. doi: 10.1016/j.mcn.2016.11.006. Epub 2016 Nov 18. Mol Cell Neurosci. 2017. PMID: 27871938 Review.
-
Neuronal, but not glial, Contactin 2 negatively regulates axon regeneration in the injured adult optic nerve.Eur J Neurosci. 2021 Mar;53(6):1705-1721. doi: 10.1111/ejn.15121. Epub 2021 Mar 18. Eur J Neurosci. 2021. PMID: 33469963
-
Structural insights into the contactin 1 - neurofascin 155 adhesion complex.Nat Commun. 2022 Nov 3;13(1):6607. doi: 10.1038/s41467-022-34302-9. Nat Commun. 2022. PMID: 36329006 Free PMC article.
References
-
- Dodd J & Jessell TM (1988) Axon guidance and the patterning of neuronal projections in vertebrates. Science 242, 692–699. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
