

BIL7 enhances plant growth by regulating the transcription factor BIL1/BZR1 during brassinosteroid signaling
- PMID: 39705127
- PMCID: PMC11771540
- DOI: 10.1111/tpj.17212
BIL7 enhances plant growth by regulating the transcription factor BIL1/BZR1 during brassinosteroid signaling
Abstract
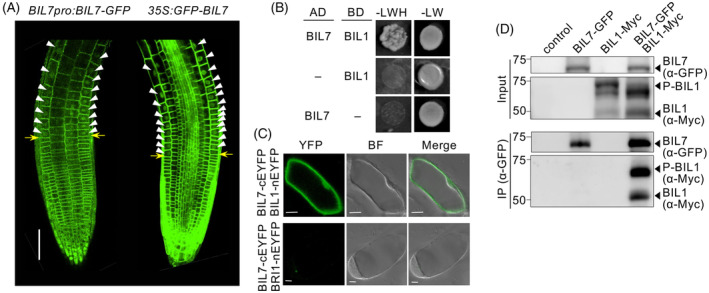
Brassinosteroids (BRs) are plant steroid hormones that regulate plant development and environmental responses. BIL1/BZR1, a master transcription factor that regulates approximately 3000 genes in the BR signaling pathway, is transported to the nucleus from the cytosol in response to BR signaling; however, the molecular mechanism underlying this process is unknown. Here, we identify a novel BR signaling factor, BIL7, that enhances plant growth and positively regulates the nuclear accumulation of BIL1/BZR1 in Arabidopsis thaliana. BIL7-overexpressing plants were resistant to the BR biosynthesis inhibitor Brz and taller than wild-type (WT) plants were due to increased cell division. BIL7 is mainly localized to the plasma membrane, but during the early stages of cell growth, it was also localized to the nucleus. BIL7 was directly phosphorylated by the kinase BIN2, and nuclear localization of BIL7 was enhanced by the BIN2 inhibitor bikinin. BIL7 was found to bind to BIL1/BZR1, and nuclear accumulation of BIL1/BZR1 was strongly enhanced by BIL7 overexpression. Finally, double overexpression of BIL1/BZR1 and BIL7 led to greatly elongated hypocotyls in the presence of Brz. These findings suggest that BIL7 mediates nuclear accumulation of BIL1/BZR1, which activates inflorescence elongation in plants via BR signaling.
Keywords: BIL1/BZR1; BIN2; NRPM; brassinosteroid; nuclear localization; plant growth; signaling pathway.
© 2024 The Author(s). The Plant Journal published by Society for Experimental Biology and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
-
- Alexandersson, E. , Saalbach, G. , Larsson, C. & Kjellbom, P. (2004) Arabidopsis plasma membrane proteomics identifies components of transport, signal transduction and membrane trafficking. Plant & Cell Physiology, 45, 1543–1556. - PubMed
-
- Asami, T. , Nakano, T. , Nakashita, H. , Sekimata, K. , Shimada, Y. & Yoshida, S. (2003) The influence of chemical genetics on plant science: shedding light on functions and mechanism of action of brassinosteroids using biosynthesis inhibitors. Journal of Plant Growth Regulation, 22, 336–349. - PubMed
-
- Asami, T. & Yoshida, S. (1999) Brassinosteroid biosynthesis inhibitors. Trends in Plant Science, 4, 348–353. - PubMed
-
- Beemster, G.T. , Fiorani, F. & Inzé, D. (2003) Cell cycle: the key to plant growth control? Trends in Plant Science, 8, 154–158. - PubMed
