

The differential interactomes of the KRAS splice variants identify BIRC6 as a ubiquitin ligase for KRAS4A
- PMID: 39705142
- PMCID: PMC12309726
- DOI: 10.1016/j.celrep.2024.115087
The differential interactomes of the KRAS splice variants identify BIRC6 as a ubiquitin ligase for KRAS4A
Abstract
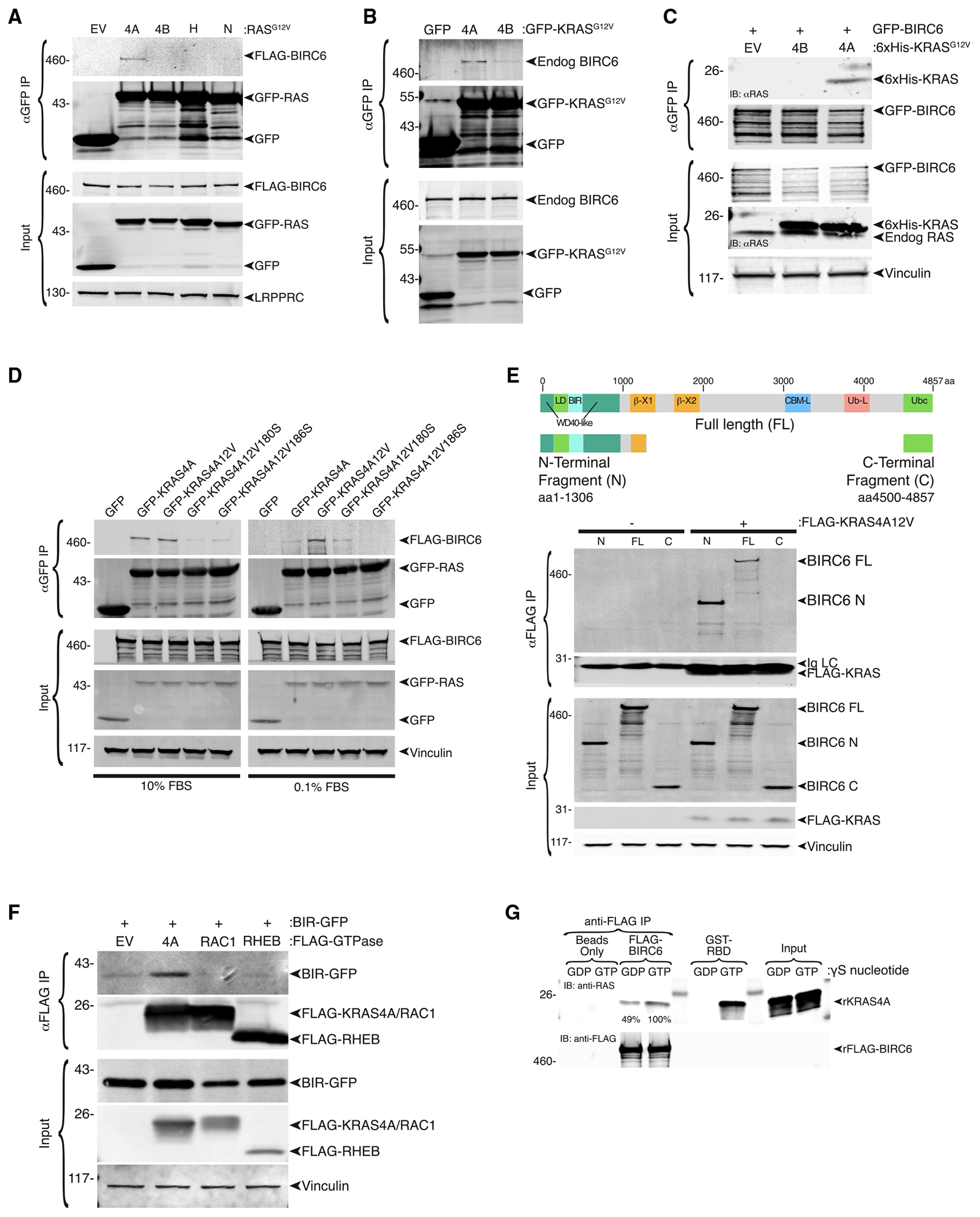
Transcripts of the KRAS locus are alternatively spliced to generate two proteins, KRAS4A and KRAS4B, which differ in their membrane-targeting sequences. These splice variants have been conserved for more than 450 million years, suggesting non-overlapping functions driven by differential membrane association. Here, we use proximity labeling to map the differential interactomes of the KRAS splice variants. We find 24 and 10 proteins that interact specifically with KRAS4A or KRAS4B, respectively. The KRAS interacting protein most specific to KRAS4A is BIRC6, a large member of the inhibitor of apoptosis protein family unique in possessing E2/E3 ubiquitin ligase activity. We find that this interaction takes place on the Golgi apparatus and results in the mono- and di-ubiquitination of KRAS4A at lysines 128 and 147. Silencing BIRC6 diminishes GTP loading of and growth stimulation by KRAS4A but not KRAS4B. Thus, BIRC6 is a ubiquitin ligase that inhibits apoptosis and also modifies KRAS4A.
Keywords: BIRC6; CP: Molecular biology; KRAS; KRAS4A; alternative splicing; inhibitor of apoptosis proteins; oncogene; post-translational modification; ubiquitination.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.S.F. is a founder, scientific advisory board (SAB) member, and equity holder of Civetta Therapeutics, Proximity Therapeutics, Stelexis Biosciences, and Neomorph, Inc. (also board of directors). He is an equity holder and SAB member for Avilar Therapeutics, Photys Therapeutics, and Ajax Therapeutics and an equity holder in Lighthorse Therapeutics and Anvia Therapeutics. E.S.F. is a consultant to Novartis, EcoR1 capital, Odyssey, and Deerfield. The Fischer lab receives or has received research funding from Deerfield, Novartis, Ajax, Interline, Bayer, and Astellas.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
