

RNA sensing induced by chromosome missegregation augments anti-tumor immunity
- PMID: 39706184
- PMCID: PMC11888943
- DOI: 10.1016/j.molcel.2024.11.025
RNA sensing induced by chromosome missegregation augments anti-tumor immunity
Abstract
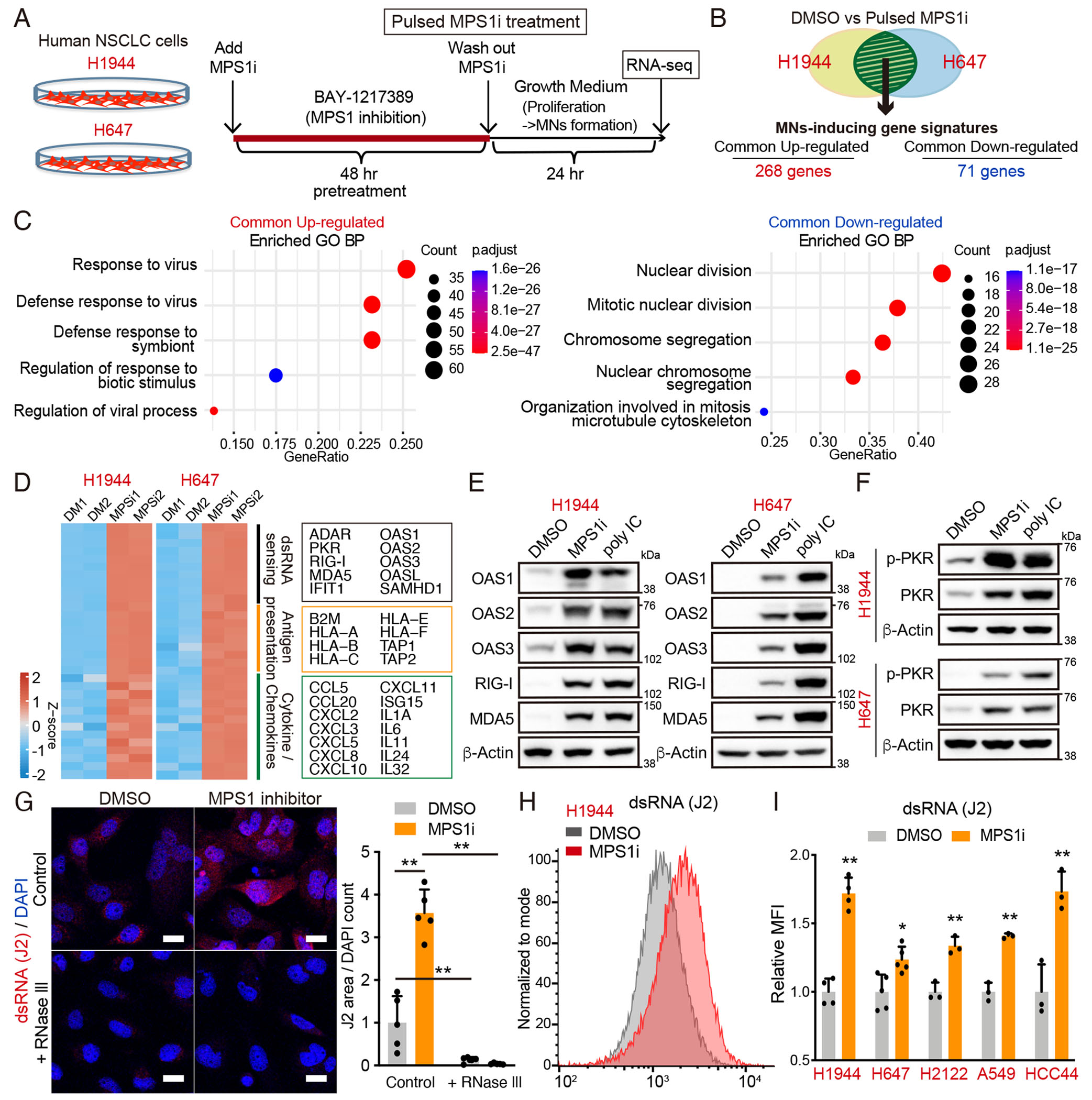
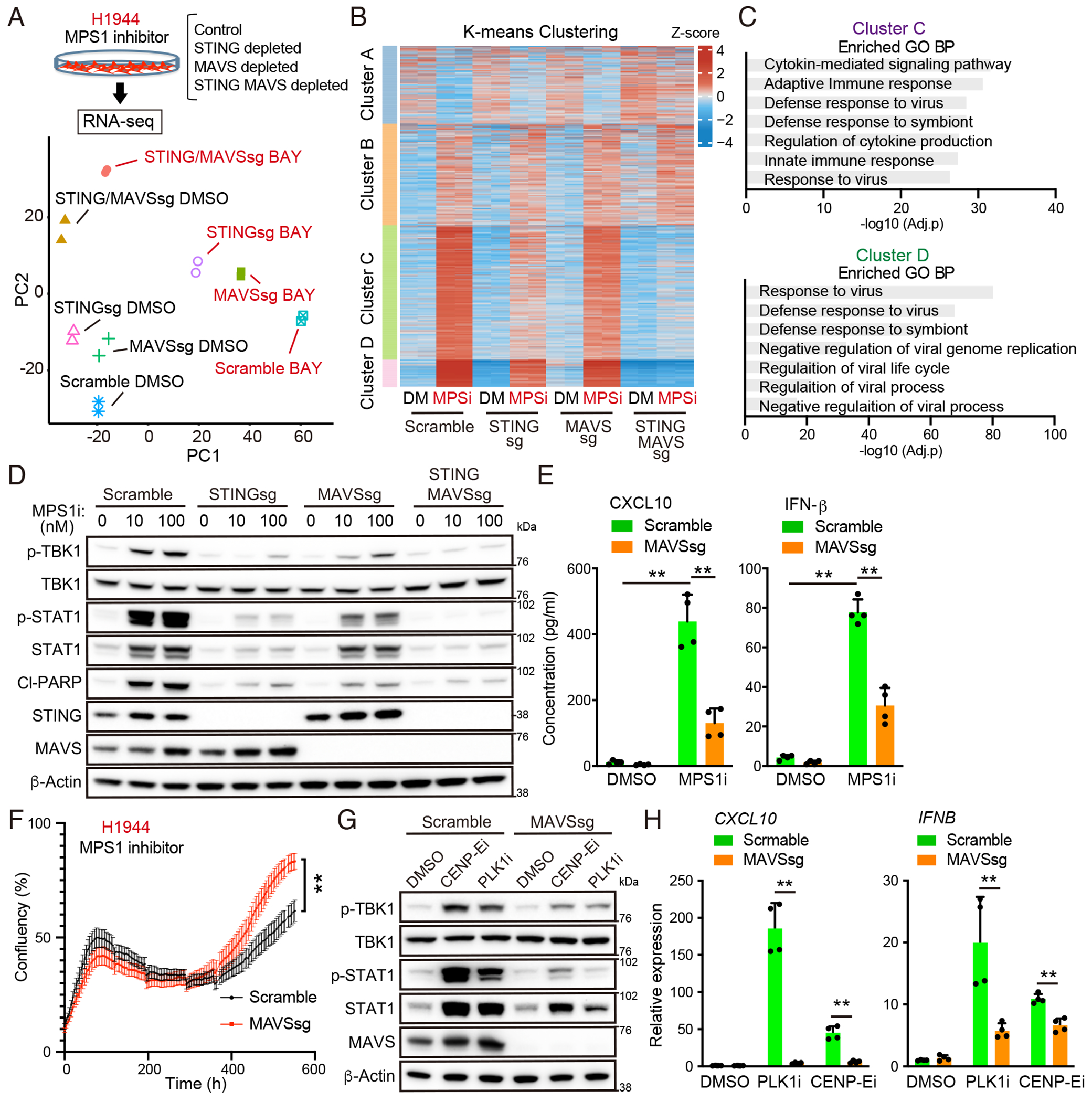
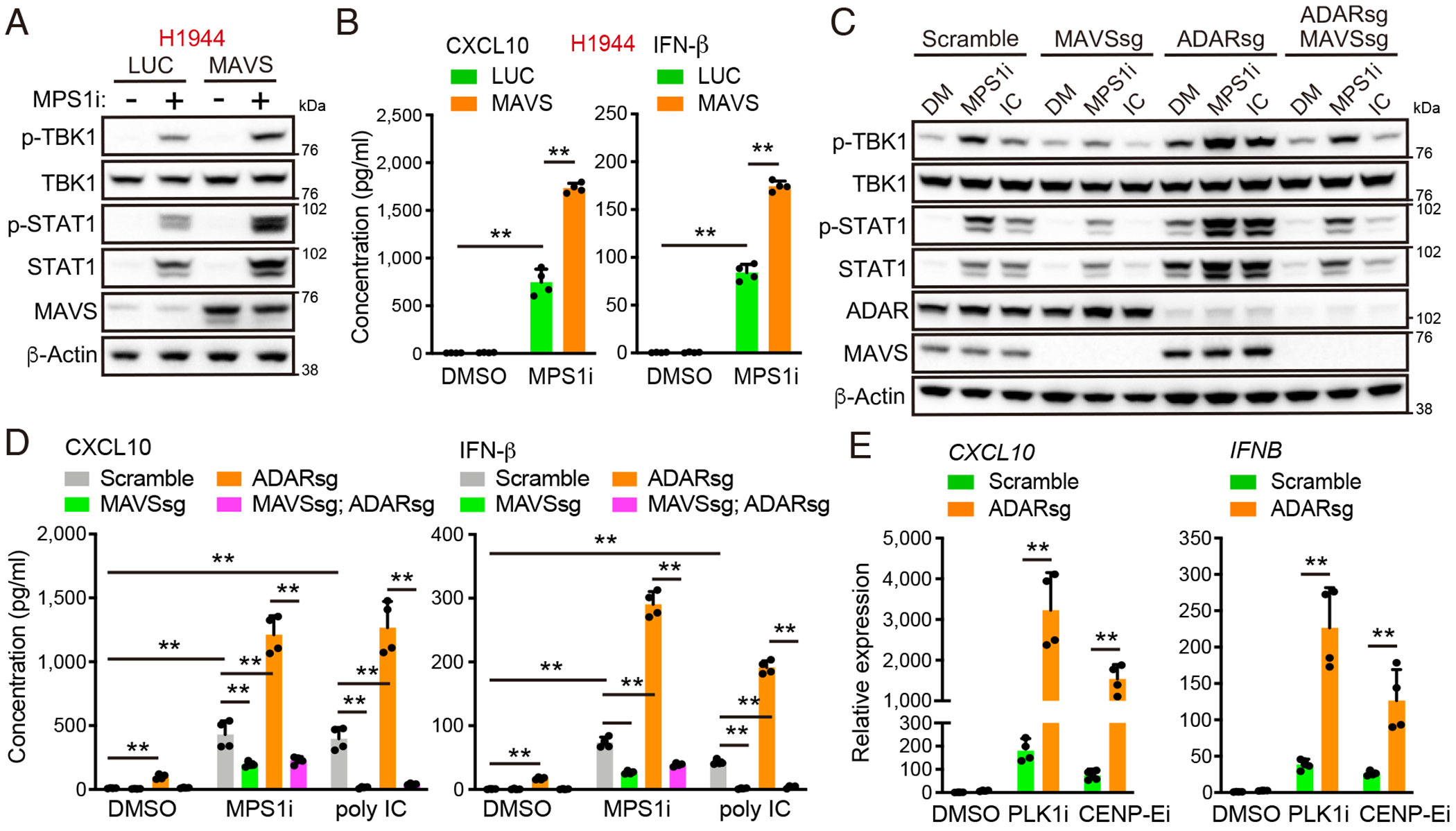
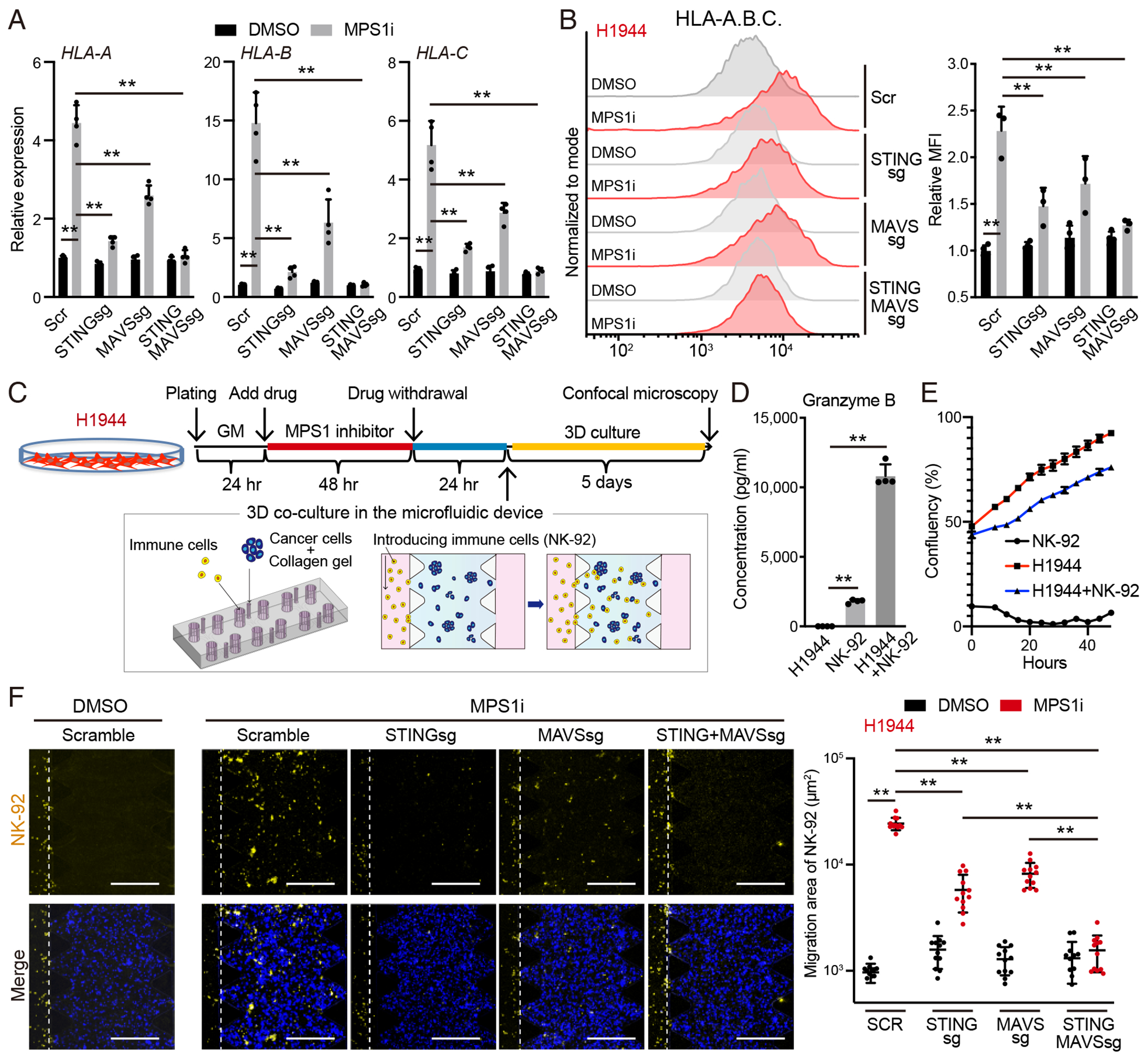
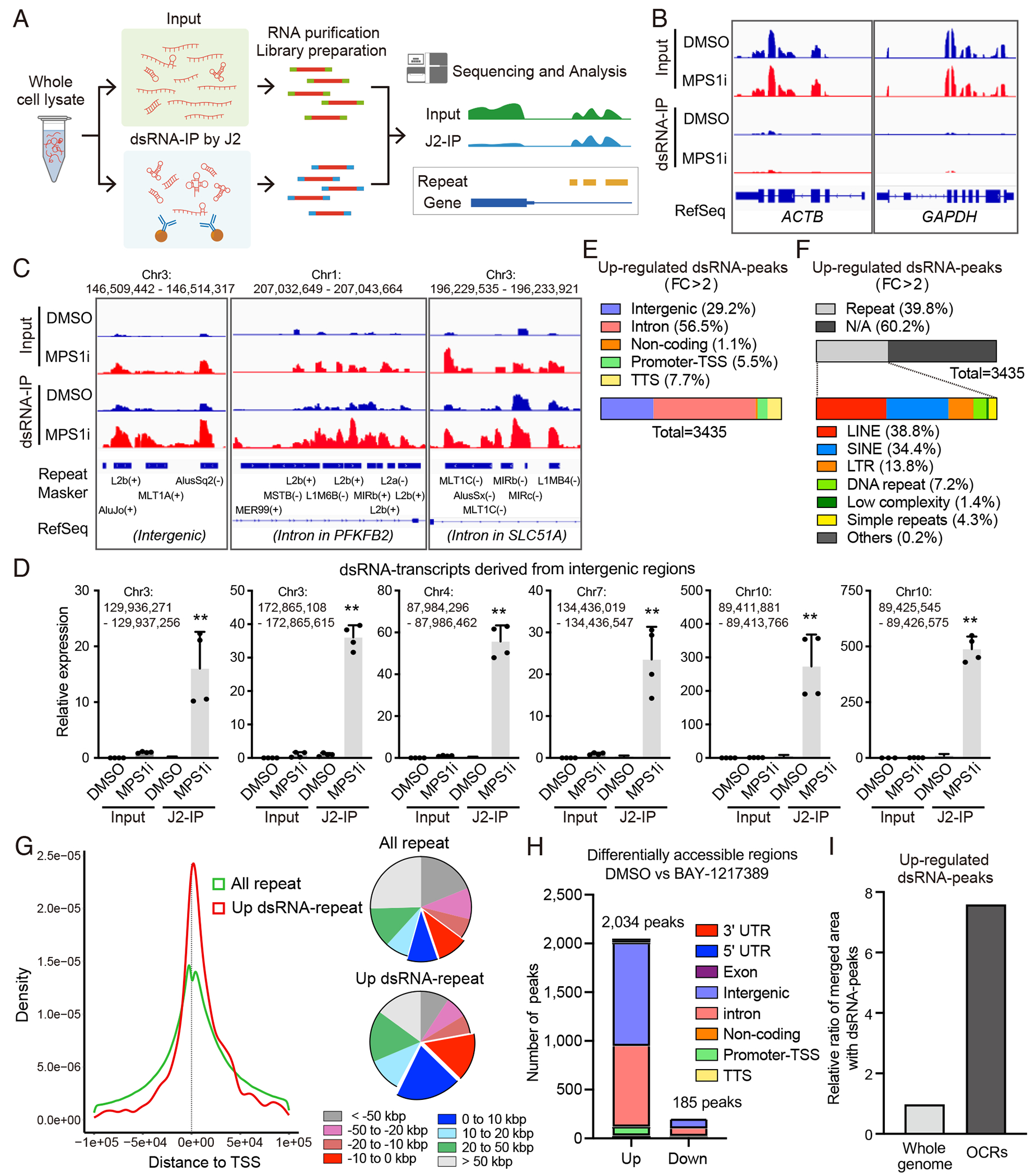
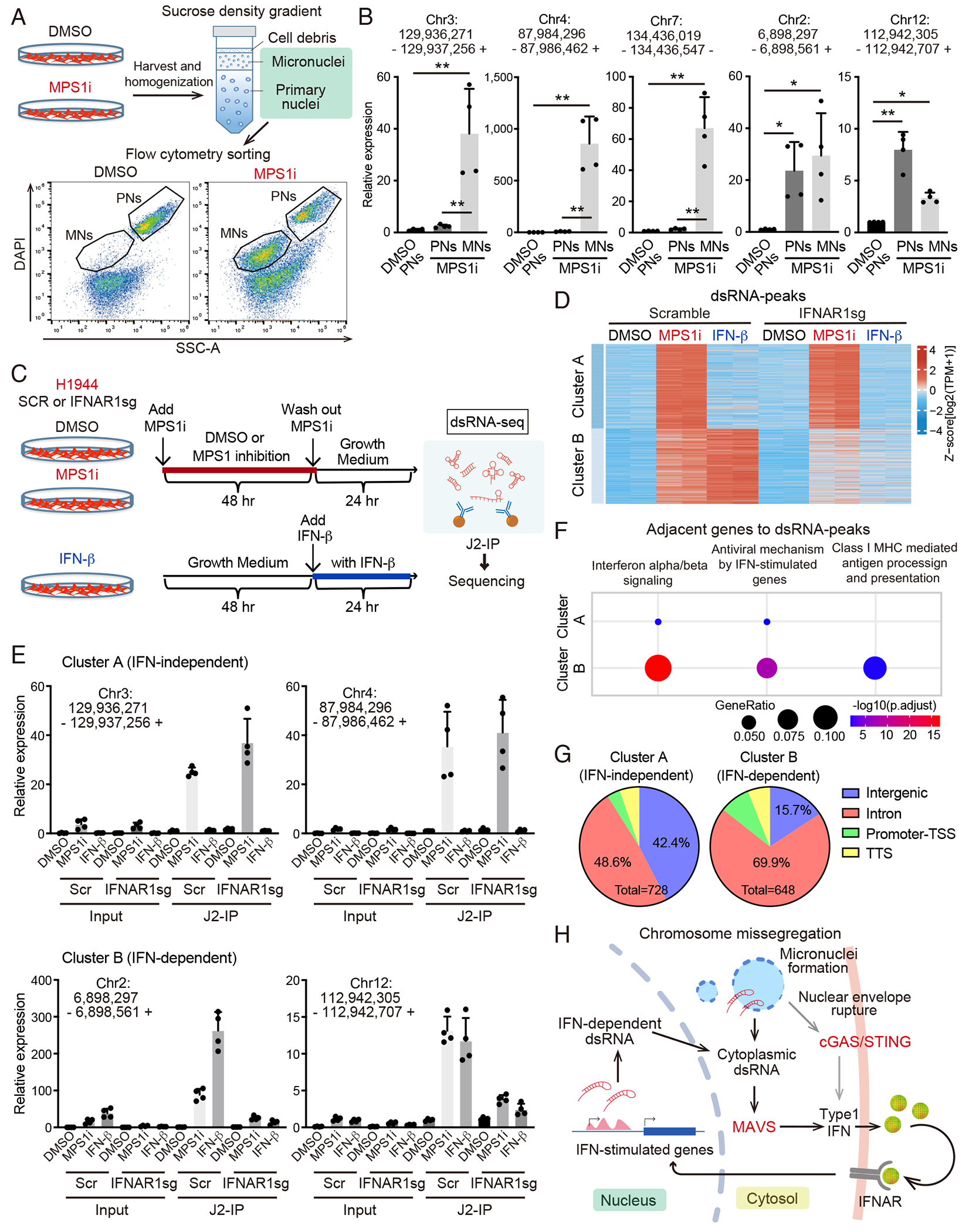
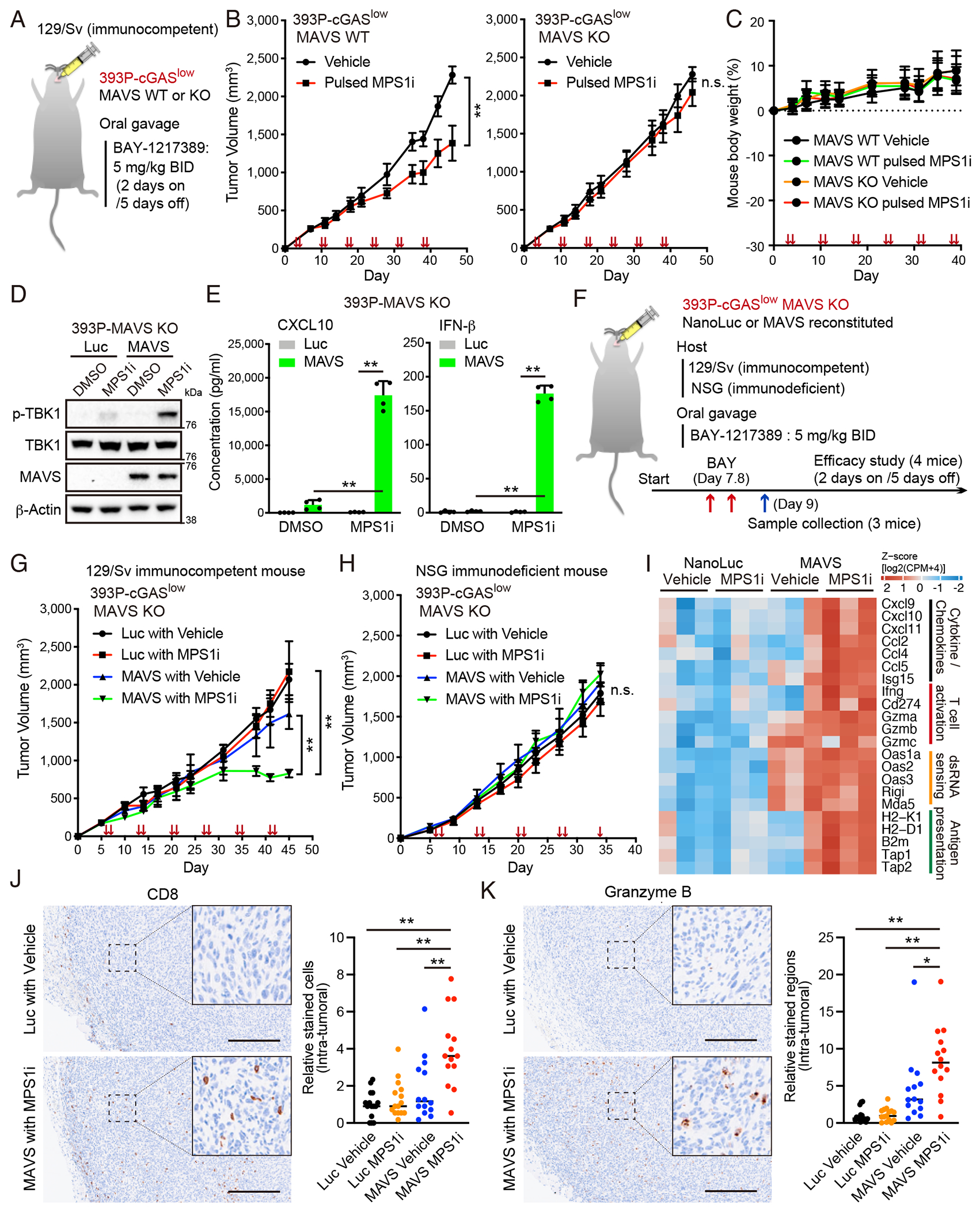
Viral mimicry driven by endogenous double-stranded RNA (dsRNA) stimulates innate and adaptive immune responses. However, the mechanisms that regulate dsRNA-forming transcripts during cancer therapy remain unclear. Here, we demonstrate that dsRNA is significantly accumulated in cancer cells following pharmacologic induction of micronuclei, stimulating mitochondrial antiviral signaling (MAVS)-mediated dsRNA sensing in conjunction with the cyclic GMP-AMP synthase (cGAS)/stimulator of interferon genes (STING) pathway. Activation of cytosolic dsRNA sensing cooperates with double-stranded DNA (dsDNA) sensing to upregulate immune cell migration and antigen-presenting machinery. Tracing of dsRNA-sequences reveals that dsRNA-forming transcripts are predominantly generated from non-exonic regions, particularly in locations proximal to genes exhibiting high chromatin accessibility. Activation of this pathway by pulsed monopolar spindle 1 (MPS1) inhibitor treatment, which potently induces micronuclei formation, upregulates cytoplasmic dsRNA sensing and thus promotes anti-tumor immunity mediated by cytotoxic lymphocyte activation in vivo. Collectively, our findings uncover a mechanism in which dsRNA sensing cooperates with dsDNA sensing to boost immune responses, offering an approach to enhance the efficacy of cancer therapies targeting genomic instability.
Keywords: MAVS; Mps1; STING; cGAS; chromosome missegregation; dsRNA; micronuclei; tumor immunity; type I interferon.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.K. has a sponsored research agreement with Boehringer Ingelheim. D.A.B. is a consultant for N of One/Qiagen and Tango Therapeutics; is a founder and shareholder in Xsphera Biosciences; has received honoraria from Merck, H3 Biomedicine/Esai, EMD Serono, Gilead Sciences, Abbvie, and Madalon Consulting; and has received research grants from BMS, Takeda, Novartis, Gilead, and Lilly.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
