

Missense mutations of the ephrin receptor EPHA1 associated with Alzheimer's disease disrupt receptor signaling functions
- PMID: 39706267
- PMCID: PMC11773478
- DOI: 10.1016/j.jbc.2024.108099
Missense mutations of the ephrin receptor EPHA1 associated with Alzheimer's disease disrupt receptor signaling functions
Abstract
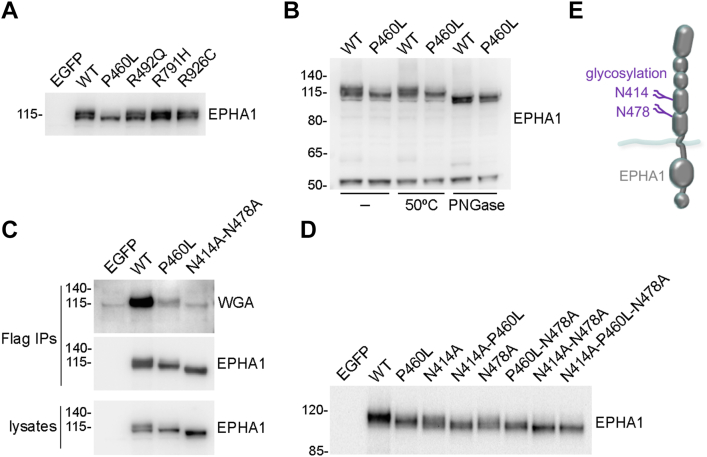
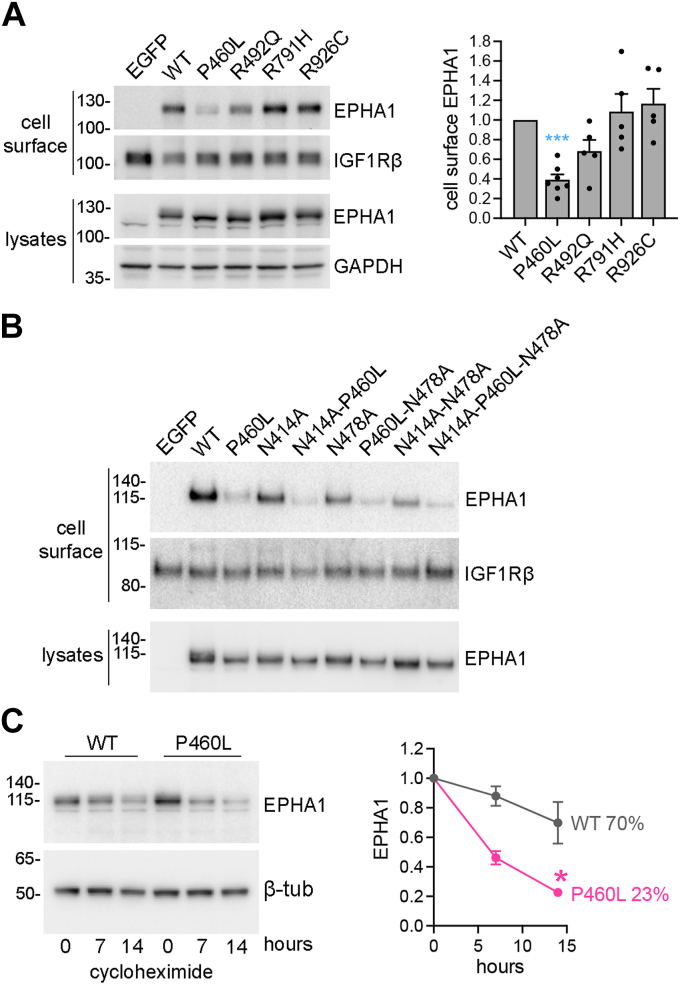
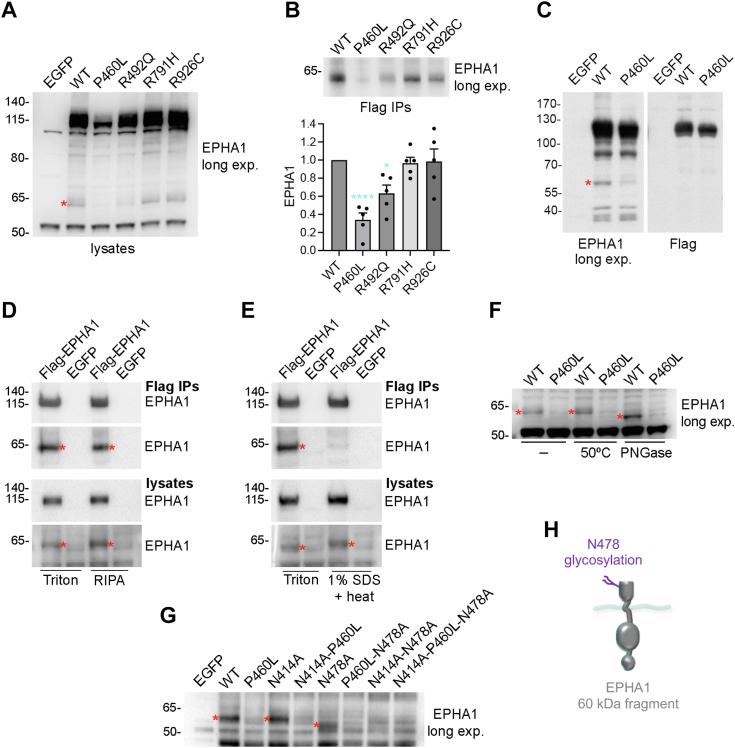
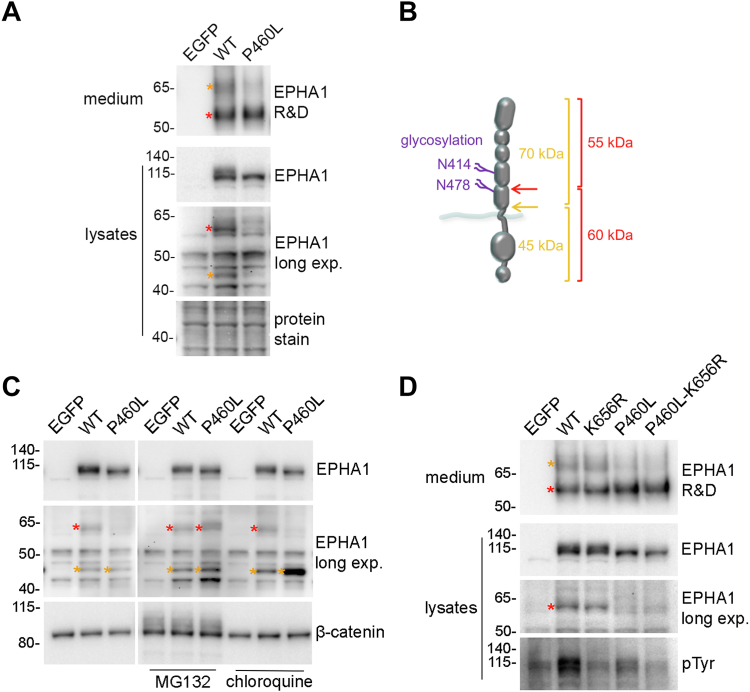
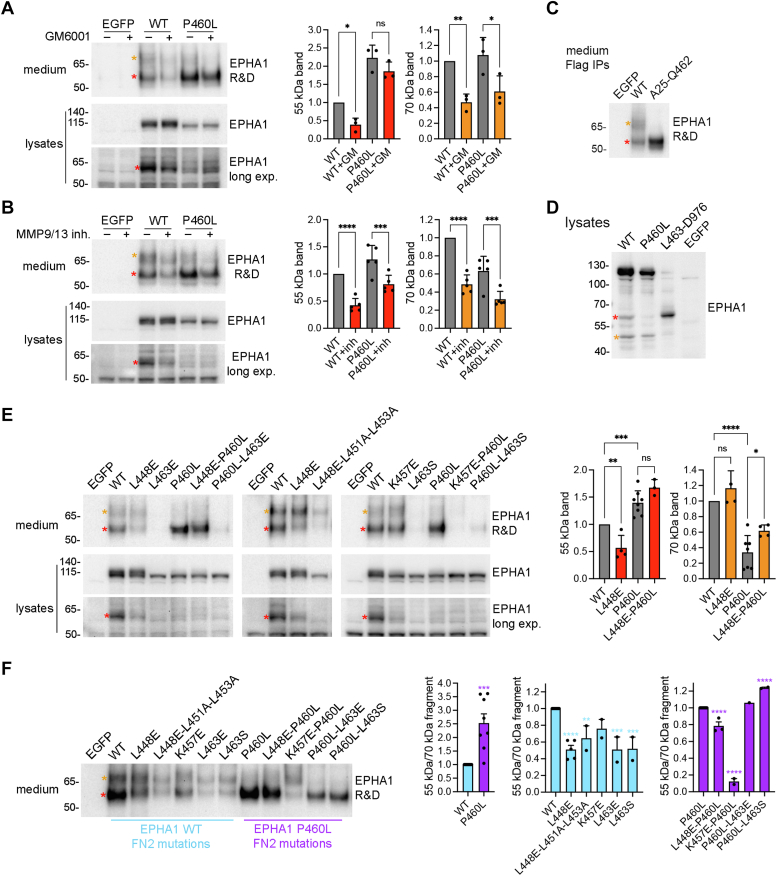
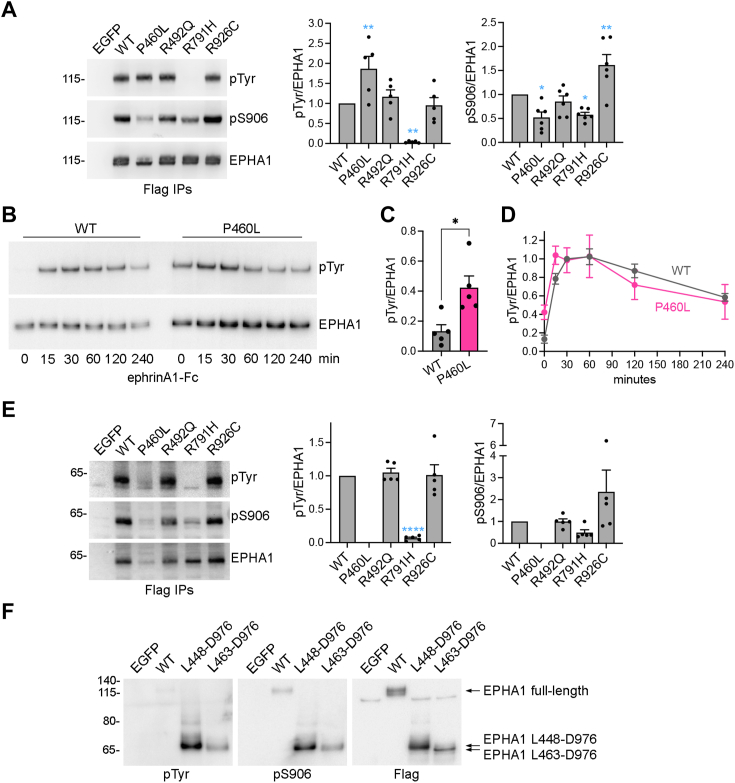
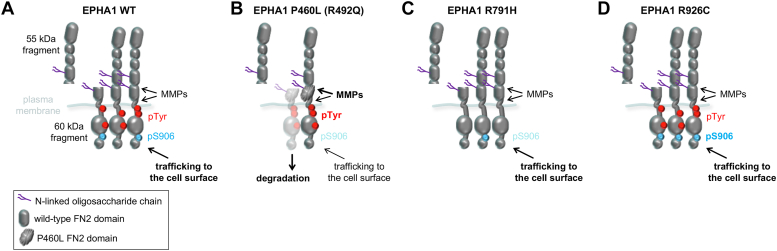
Missense mutations in the EPHA1 receptor tyrosine kinase have been identified in Alzheimer's patients. To gain insight into their potential role in disease pathogenesis, we investigated the effects of four of these mutations. We show that the P460L mutation in the second fibronectin type III (FN2) domain drastically reduces EPHA1 cell surface localization while increasing tyrosine phosphorylation of the cell surface-localized receptor. The R791H mutation in the kinase domain abolishes EPHA1 tyrosine phosphorylation, indicating abrogation of kinase-dependent signaling. Furthermore, both mutations decrease EPHA1 phosphorylation on S906 in the kinase-SAM linker region, suggesting impairment of a noncanonical form of signaling regulated by serine/threonine kinases. The R492Q mutation, also in the FN2 domain, has milder effects than the P460L mutation while the R926C mutation in the SAM domain increases S906 phosphorylation. We also found that EPHA1 undergoes constitutive proteolytic cleavage in the FN2 domain, generating a soluble 55 kDa N-terminal fragment containing the ligand-binding domain and a transmembrane 60 kDa C-terminal fragment. The 60 kDa WT fragment is phosphorylated on both tyrosine residues and S906, suggesting signaling functions. The P460L mutant 60 kDa fragment undergoes proteasomal degradation and the R791H mutant fragment lacks tyrosine phosphorylation and has decreased S906 phosphorylation. These findings advance our understanding of EPHA1 signaling mechanisms and support the notion that alterations in EPHA1 signaling due to missense mutations contribute to Alzheimer's disease pathogenesis.
Keywords: Alzheimer’s disease; Eph receptor; N-linked glycosylation; phosphoserine; phosphotyrosine; proteolytic cleavage; receptor modification; receptor tyrosine kinase.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
