

HSP60 controls mitochondrial ATP generation for optimal virus-specific IL-21-producing CD4 and cytotoxic CD8 memory T cell responses
- PMID: 39709477
- PMCID: PMC11663223
- DOI: 10.1038/s42003-024-07326-8
HSP60 controls mitochondrial ATP generation for optimal virus-specific IL-21-producing CD4 and cytotoxic CD8 memory T cell responses
Abstract
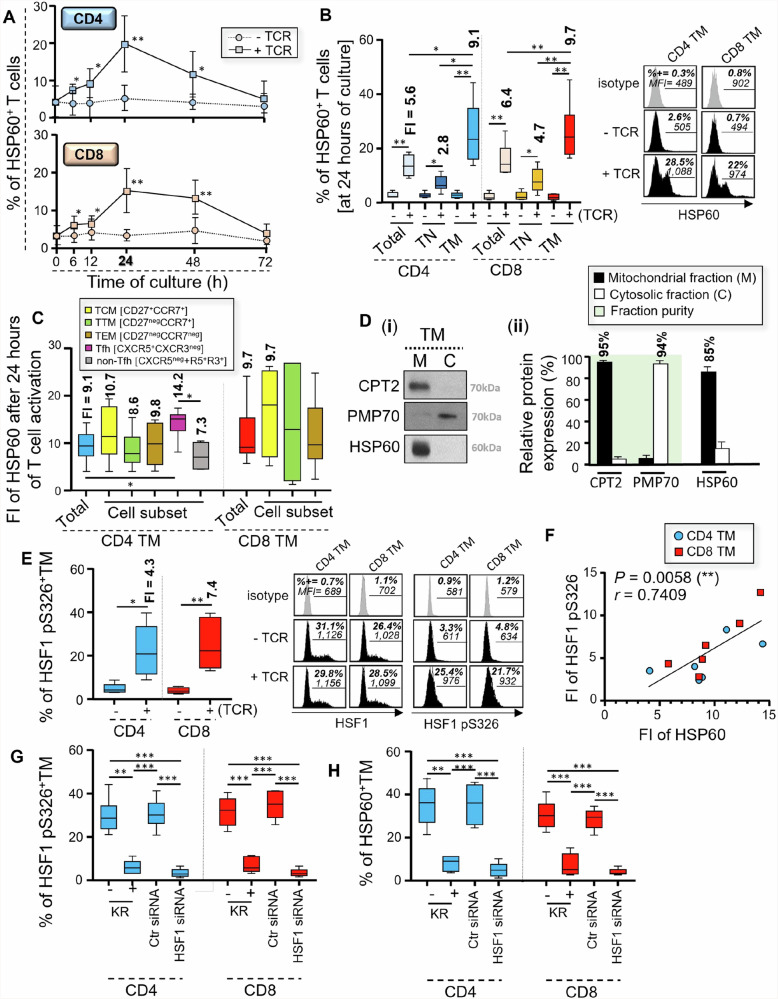
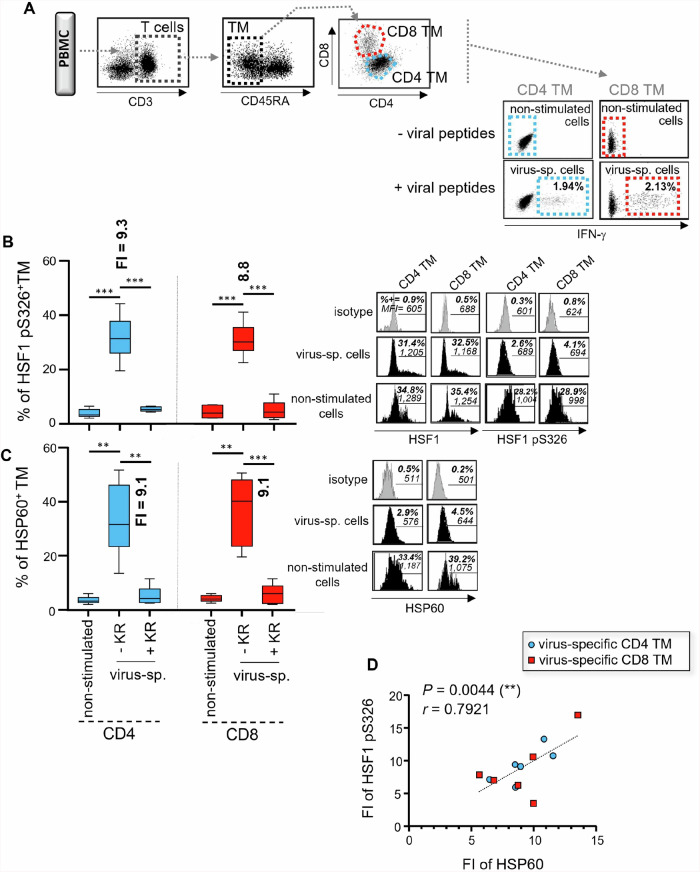
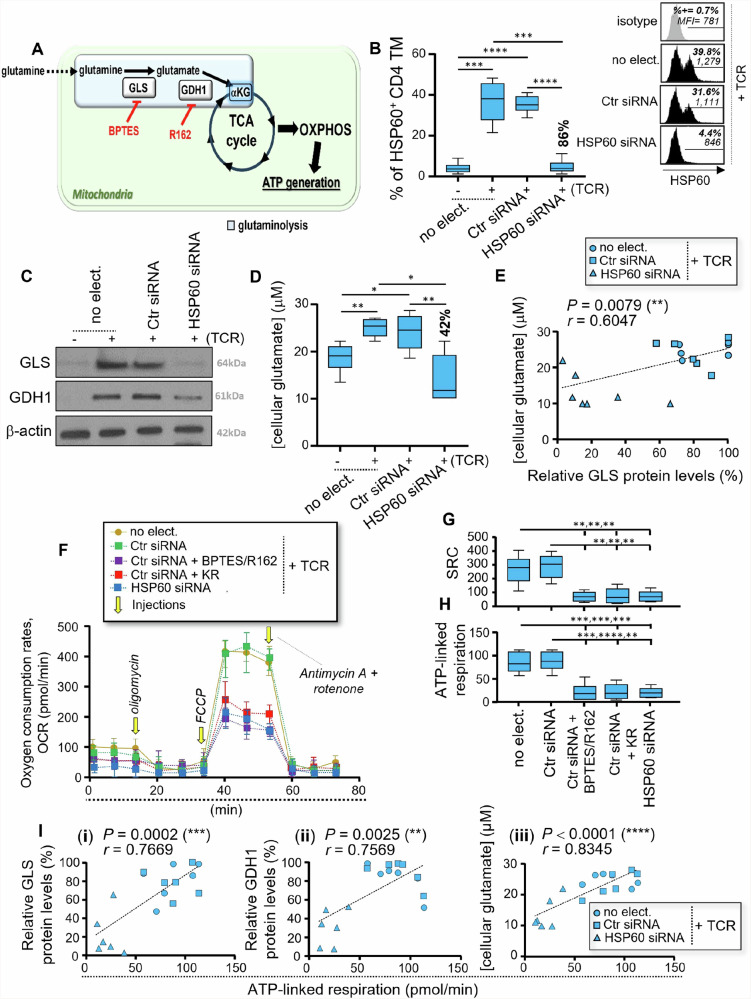
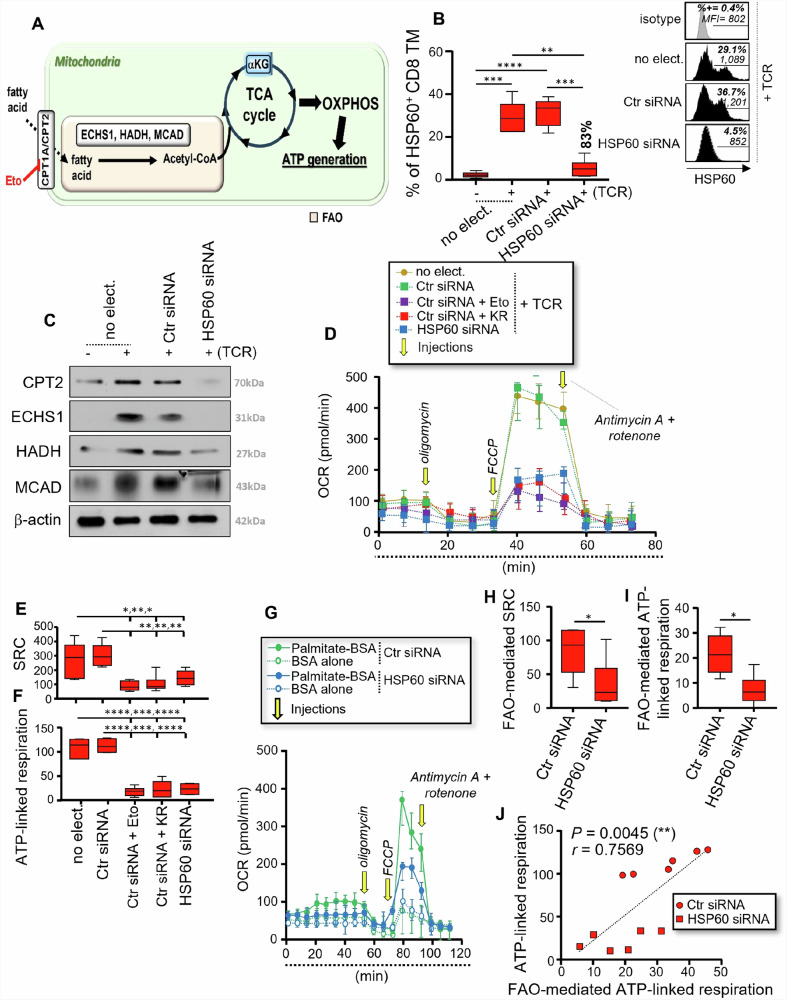
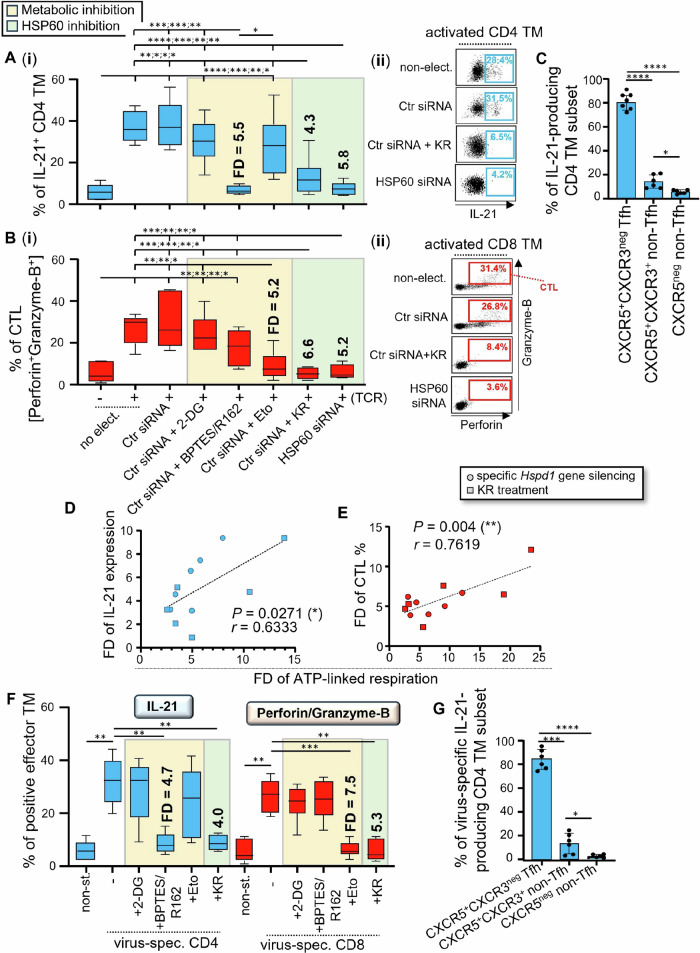
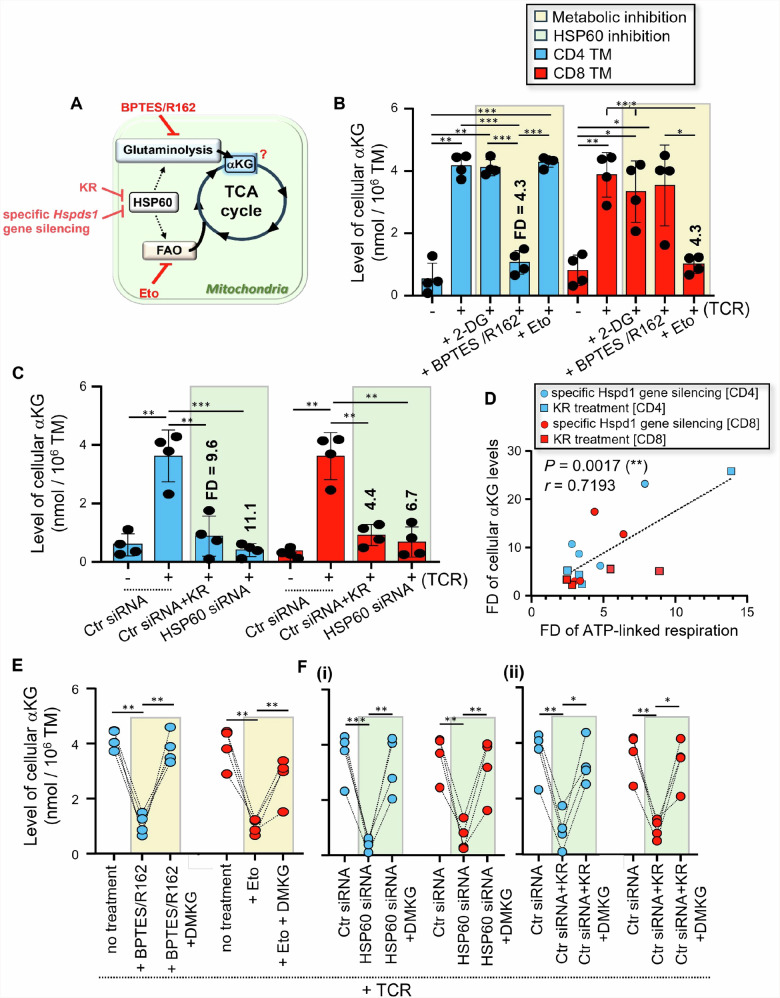
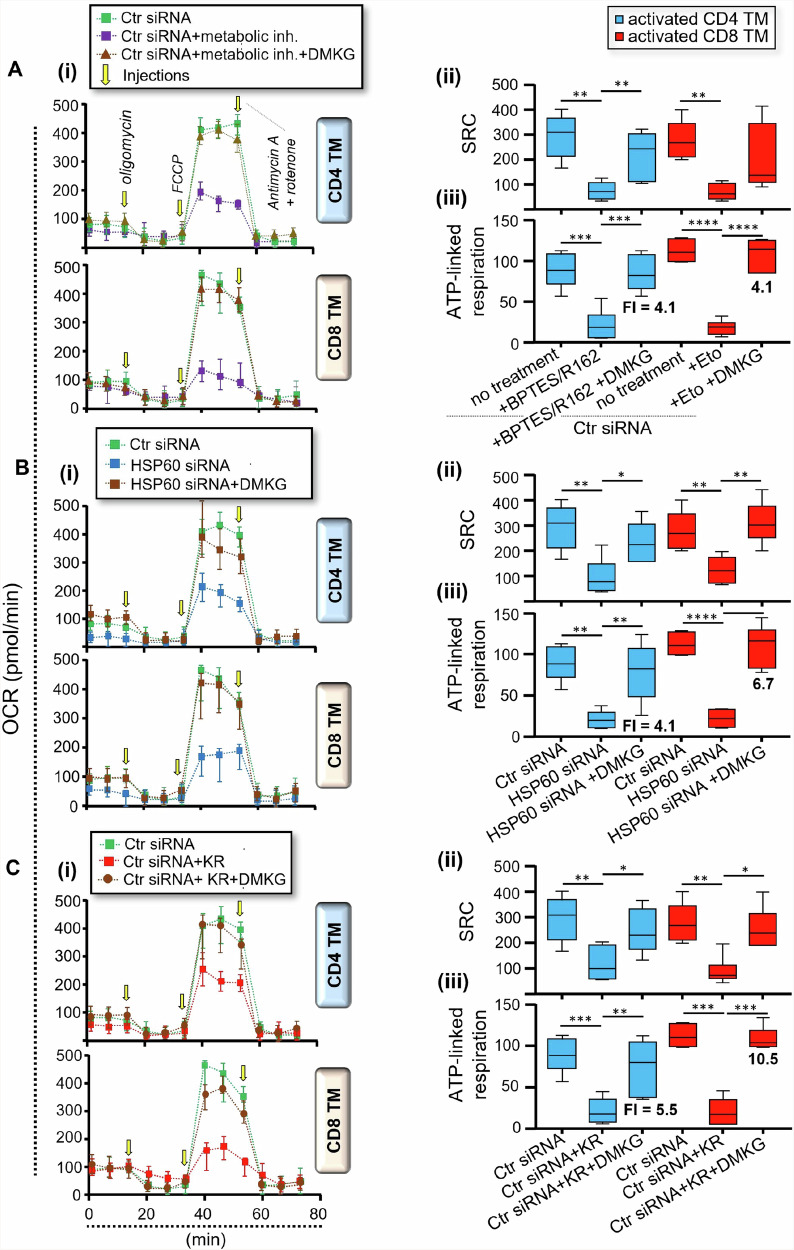
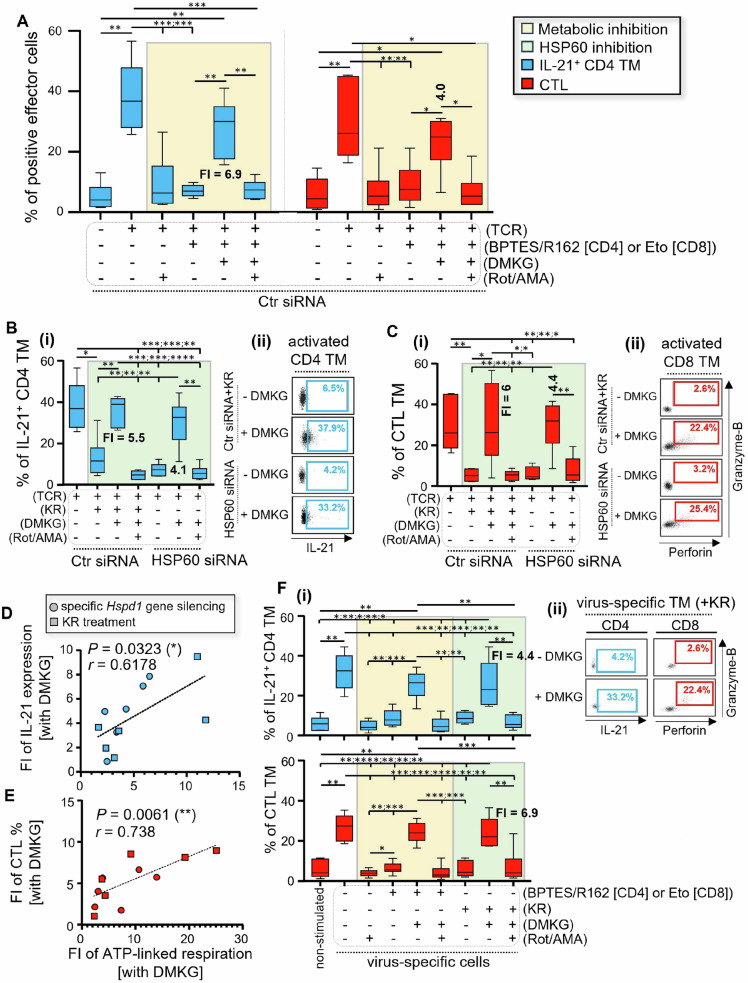
We have shown that virus-specific CD4 and CD8 memory T cells (TM) induce autophagy after T cell receptor (TCR) engagement to provide free glutamine and fatty acids, including in people living with HIV-1 (PLWH). These nutrients fuel mitochondrial ATP generation through glutaminolysis and fatty acid oxidation (FAO) pathways, to fulfill the bioenergetic demands for optimal IL-21 and cytotoxic molecule production in CD4 and CD8 cells, respectively. Here, we expand our knowledge on how the metabolic events that occur in the mitochondria of virus-specific TM down-stream of the autophagy are regulated. We show that HSP60 chaperone positively regulates the protein levels for multiple glutaminolysis- and FAO-related enzymes, thereby actively fueling the levels of cellular alpha-ketoglutarate (αKG) and related mitochondrial ATP-dependent antiviral T cell immunity in both CD4 and CD8 TM. Finally, we provide a way to rescue defective ATP generation in mitochondria and dependent effector functions in virus-specific TM including anti-HIV-1 protective responses, when HSP60 expression is impaired after TCR engagement in patients, in the form of dimethyl 2-oxoglutarate (DMKG) supplementation.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
- RGPIN-2018-05272/Canadian Network for Research and Innovation in Machining Technology, Natural Sciences and Engineering Research Council of Canada (NSERC Canadian Network for Research and Innovation in Machining Technology)
- 486498/Gouvernement du Canada | Instituts de Recherche en Santé du Canada | CIHR Skin Research Training Centre (Skin Research Training Centre)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
