

Embracing native diversity to enhance the maximum quantum efficiency of photosystem II in maize
- PMID: 39711175
- PMCID: PMC11702984
- DOI: 10.1093/plphys/kiae670
Embracing native diversity to enhance the maximum quantum efficiency of photosystem II in maize
Abstract
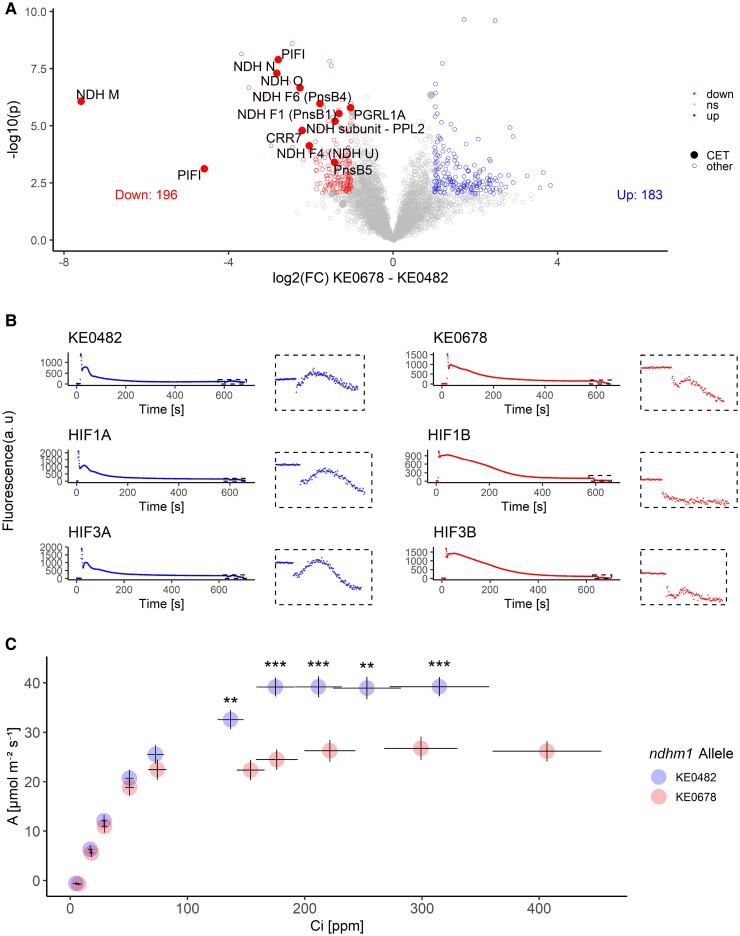
The sustainability of maize cultivation would benefit tremendously from early sowing, but is hampered by low temperatures during early development in temperate climates. We show that allelic variation within the gene encoding subunit M of the NADH-dehydrogenase-like (NDH) complex (ndhm1) in a European maize landrace affects several quantitative traits that are relevant during early development in cold climates through NDH-mediated cyclic electron transport around photosystem I, a process crucial for photosynthesis and photoprotection. Beginning with a genome-wide association study for maximum potential quantum yield of photosystem II in dark-adapted leaves (Fv/Fm), we capitalized on the large phenotypic effects of a hAT transposon insertion in ndhm1 on multiple quantitative traits (early plant height [EPH], Fv/Fm, chlorophyll content, and cold tolerance) caused by the reduced protein levels of NDHM and associated NDH components. Analysis of the ndhm1 native allelic series revealed a rare allele of ndhm1 that is associated with small albeit significant improvements of Fv/Fm, photosystem II efficiency in light-adapted leaves (ΦPSII), and EPH compared with common alleles. Our work showcases the extraction of favorable alleles from locally adapted landraces, offering an efficient strategy for broadening the genetic variation of elite germplasm by breeding or genome editing.
© The Author(s) 2024. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures






Similar articles
-
Dissecting the genetics of cold tolerance in a multiparental maize population.Theor Appl Genet. 2020 Feb;133(2):503-516. doi: 10.1007/s00122-019-03482-2. Epub 2019 Nov 18. Theor Appl Genet. 2020. PMID: 31740990
-
Studies of the Ndh complex and photosystem II from mesophyll and bundle sheath chloroplasts of the C4-type plant Zea mays.J Plant Physiol. 2006 Jul;163(8):800-8. doi: 10.1016/j.jplph.2005.11.014. Epub 2006 Jan 24. J Plant Physiol. 2006. PMID: 16436305
-
Cold stress effects on PSI photochemistry in Zea mays: differential increase of FQR-dependent cyclic electron flow and functional implications.Plant Cell Physiol. 2011 Jun;52(6):1042-54. doi: 10.1093/pcp/pcr056. Epub 2011 May 4. Plant Cell Physiol. 2011. PMID: 21546369
-
Physiological, Photosynthetic Characteristic and Transcriptome Analysis of PsnWRKY70 Transgenic Populus simonii × Populus nigra Under Salt Stress.Int J Mol Sci. 2024 Dec 25;26(1):81. doi: 10.3390/ijms26010081. Int J Mol Sci. 2024. PMID: 39795940 Free PMC article.
-
Diversity in global maize germplasm: characterization and utilization.J Biosci. 2012 Nov;37(5):843-55. doi: 10.1007/s12038-012-9227-1. J Biosci. 2012. PMID: 23107920 Review.
Cited by
-
Rapid cycling genomic selection in maize landraces.Theor Appl Genet. 2025 Mar 17;138(4):75. doi: 10.1007/s00122-025-04855-6. Theor Appl Genet. 2025. PMID: 40094967 Free PMC article.
-
Designing a nitrogen-efficient cold-tolerant maize for modern agricultural systems.Plant Cell. 2025 Jul 1;37(7):koaf139. doi: 10.1093/plcell/koaf139. Plant Cell. 2025. PMID: 40673831 Free PMC article. Review.
References
-
- Asada K, Heber U, Schreiber U. Electron flow to the intersystem chain from stromal components and cyclic electron flow in maize chloroplasts, as detected in intact leaves by monitoring redox change of P700 and chlorophyll fluorescence. Plant Cell Physiol. 1993:34:39–50. 10.1093/oxfordjournals.pcp.a078398 - DOI
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol. 1995:57(1):289–300. 10.1111/j.2517-6161.1995.tb02031.x - DOI
-
- Blankenagel S, Eggels S, Frey M, Grill E, Bauer E, Dawid C, Fernie AR, Haberer G, Hammerl R, Barbosa Medeiros D, et al. Natural alleles of the abscisic acid catabolism gene ZmAbh4 modulate water use efficiency and carbon isotope discrimination in maize. Plant Cell. 2022:34(10):3860–3872. 10.1093/plcell/koac200 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
