

This is a preprint.
Covalent-fragment screening identifies selective inhibitors of multiple Staphylococcus aureus serine hydrolases important for growth and biofilm formation
- PMID: 39711551
- PMCID: PMC11661381
- DOI: 10.21203/rs.3.rs-5494070/v1
Covalent-fragment screening identifies selective inhibitors of multiple Staphylococcus aureus serine hydrolases important for growth and biofilm formation
Update in
-
Identification of covalent inhibitors of Staphylococcus aureus serine hydrolases important for virulence and biofilm formation.Nat Commun. 2025 May 30;16(1):5046. doi: 10.1038/s41467-025-60367-3. Nat Commun. 2025. PMID: 40447595 Free PMC article.
Abstract
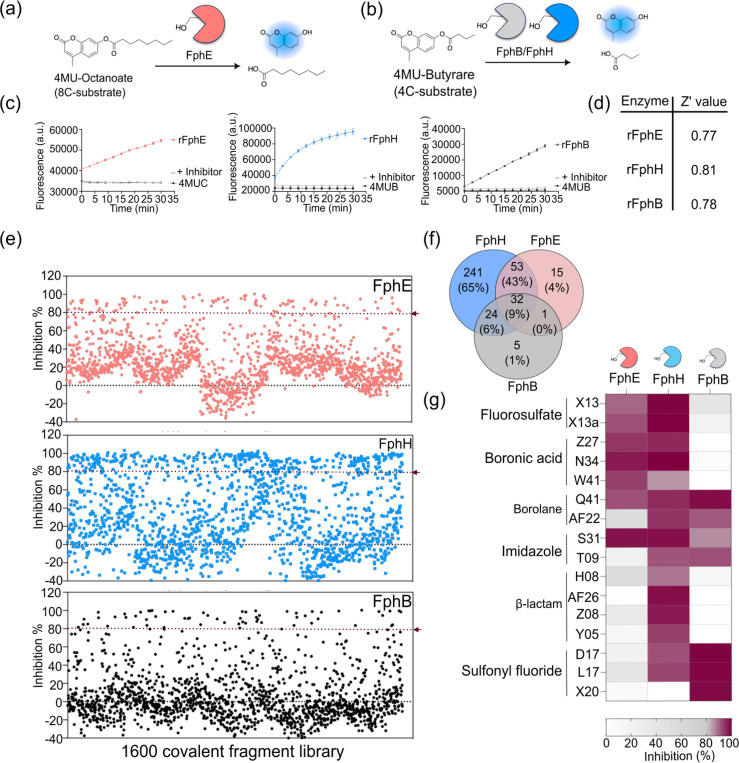
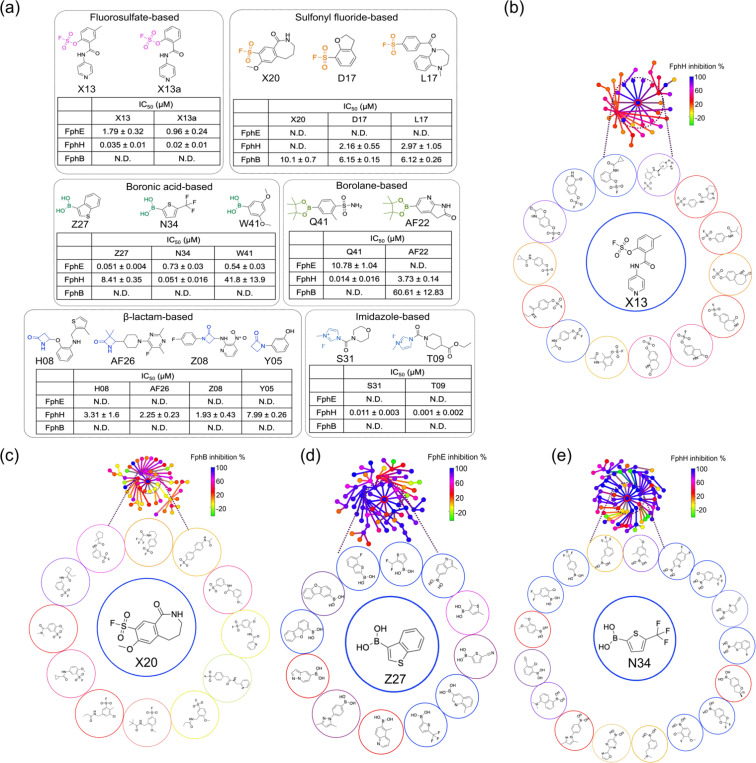
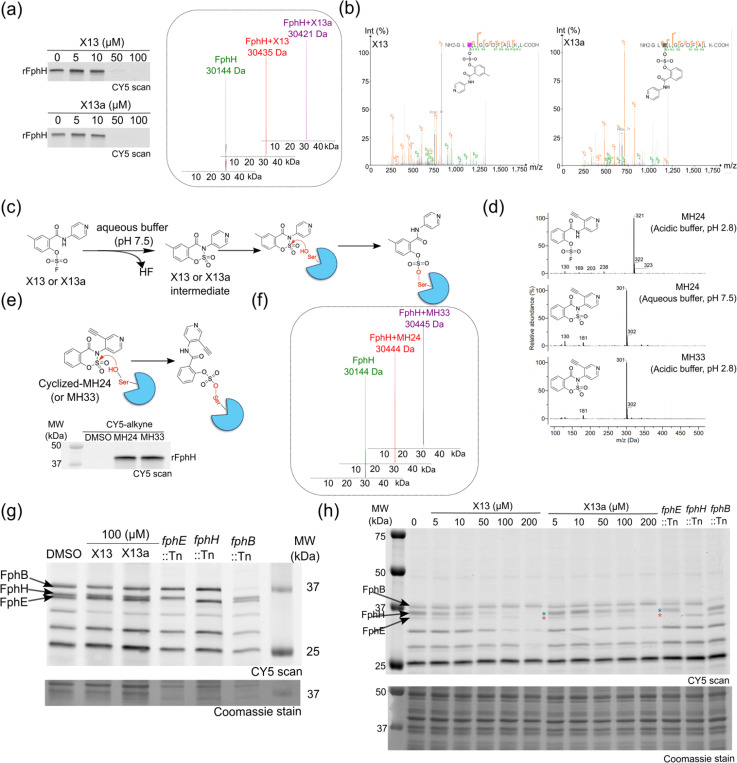
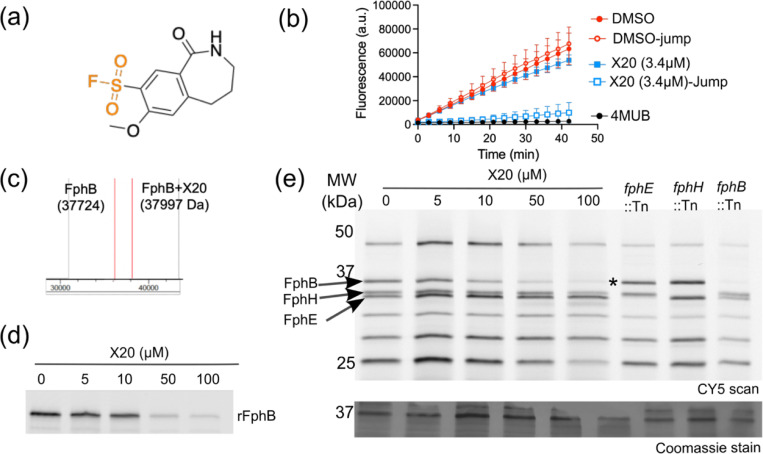
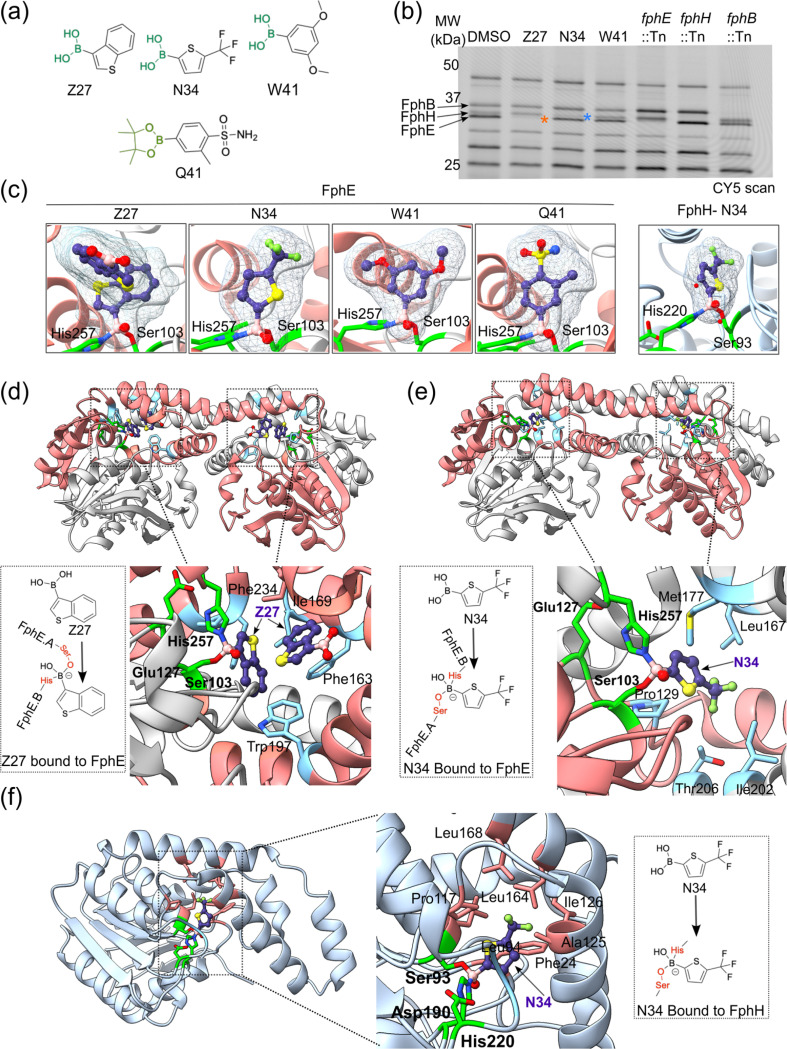
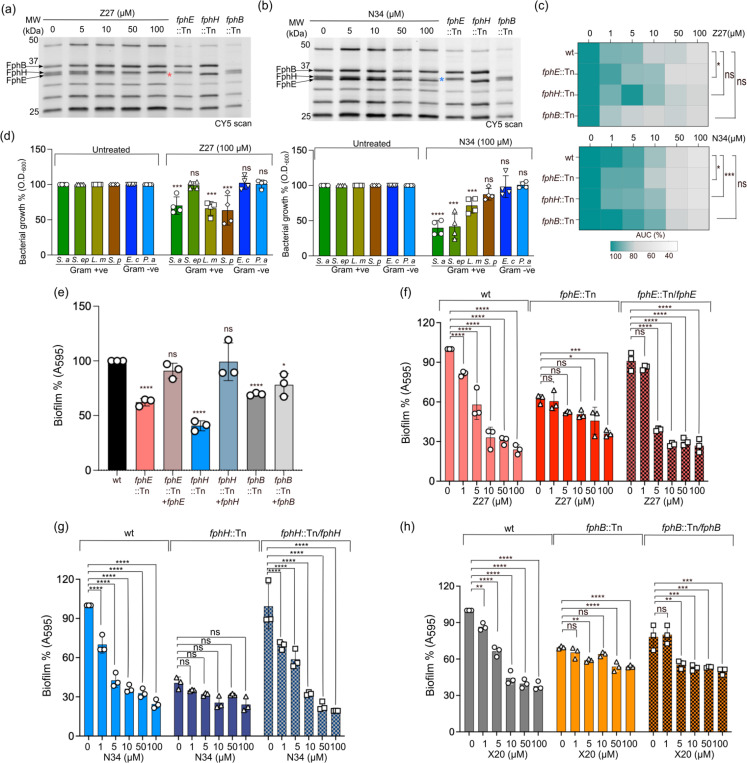
Staphylococcus aureus is a leading cause of bacteria-associated mortality worldwide. This is largely because infection sites are often difficult to localize and the bacteria forms biofilms which are not effectively cleared using classical antibiotics. Therefore, there is a need for new tools to both image and treat S. aureus infections. We previously identified a group of S. aureus serine hydrolases known as fluorophosphonate-binding hydrolases (Fphs), which regulate aspects of virulence and lipid metabolism. However, because their structures are similar and their functions overlap, it remains challenging to distinguish the specific roles of individual members of this family. In this study, we applied a high-throughput screening approach using a library of covalent electrophiles to identify inhibitors for FphB, FphE, and FphH. We identified inhibitors that irreversibly bind to the active-site serine residue of each enzyme with high potency and selectivity without requiring extensive medicinal chemistry optimization. Structural and biochemical analysis identified novel binding modes for several of the inhibitors. Selective inhibitors of FphH impaired both bacterial growth and biofilm formation while Inhibitors of FphB and FphE had no impact on cell growth and only limited impact on biofilm formation. These results suggest that all three hydrolases likely play functional, but non-equivalent roles in biofilm formation and FphH is a potential target for development of therapeutics that have both antibiotic and anti-biofilm activity. Overall, we demonstrate that focused covalent fragment screening can be used to rapidly identify highly potent and selective electrophiles targeting bacterial serine hydrolases. This approach could be applied to other classes of lipid hydrolases in diverse pathogens or higher eukaryotes.
Figures
Similar articles
-
Identification of covalent inhibitors of Staphylococcus aureus serine hydrolases important for virulence and biofilm formation.Nat Commun. 2025 May 30;16(1):5046. doi: 10.1038/s41467-025-60367-3. Nat Commun. 2025. PMID: 40447595 Free PMC article.
-
An mRNA Display Approach for Covalent Targeting of a Staphylococcus aureus Virulence Factor.bioRxiv [Preprint]. 2024 Nov 8:2024.11.06.622387. doi: 10.1101/2024.11.06.622387. bioRxiv. 2024. Update in: J Am Chem Soc. 2025 Mar 12;147(10):8312-8325. doi: 10.1021/jacs.4c15713. PMID: 39574702 Free PMC article. Updated. Preprint.
-
Biochemical and Cellular Characterization of the Function of Fluorophosphonate-Binding Hydrolase H (FphH) in Staphylococcus aureus Support a Role in Bacterial Stress Response.ACS Infect Dis. 2023 Nov 10;9(11):2119-2132. doi: 10.1021/acsinfecdis.3c00246. Epub 2023 Oct 12. ACS Infect Dis. 2023. PMID: 37824340 Free PMC article.
-
Beyond the double helix: the multifaceted landscape of extracellular DNA in Staphylococcus aureus biofilms.Front Cell Infect Microbiol. 2024 Jun 5;14:1400648. doi: 10.3389/fcimb.2024.1400648. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38903938 Free PMC article. Review.
-
Strategies for Tuning the Selectivity of Chemical Probes that Target Serine Hydrolases.Cell Chem Biol. 2020 Aug 20;27(8):937-952. doi: 10.1016/j.chembiol.2020.07.008. Epub 2020 Jul 28. Cell Chem Biol. 2020. PMID: 32726586 Free PMC article. Review.
References
-
- Lee A. S. et al. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primer 4, 1–23 (2018). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
