

Enhancing aphid resistance in horticultural crops: a breeding prospective
- PMID: 39712868
- PMCID: PMC11659385
- DOI: 10.1093/hr/uhae275
Enhancing aphid resistance in horticultural crops: a breeding prospective
Abstract
Increasing agricultural losses caused by insect infestations are a significant problem, so it is important to generate pest-resistant crop varieties to address this issue. Several reviews have examined aphid-plant interactions from an entomological perspective. However, few have specifically focused on plant resistance mechanisms to aphids and their applications in breeding for aphid resistance. In this review, we first outline the types of resistance to aphids in plants, namely antixenosis, tolerance (cell wall lignification, resistance proteins), and antibiosis, and we discuss strategies based on each of these resistance mechanisms to generate plant varieties with improved resistance. We then outline research on the complex interactions amongst plants, viruses, and aphids, and discuss how aspects of these interactions can be exploited to improve aphid resistance. A deeper understanding of the epigenetic mechanisms related to induced resistance, i.e. the phenomenon where plants become more resistant to a stress they have encountered previously, may allow for its exploitation in breeding for aphid resistance. Wild relatives of crop plants serve as important sources of resistance traits. Genes related to these traits can be introduced into cultivated crop varieties by breeding or genetic modification, and de novo domestication of wild varieties can be used to exploit multiple excellent characteristics, including aphid resistance. Finally, we discuss the use of molecular design breeding, genomic data, and gene editing to generate new aphid-resistant, high-quality crop varieties.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nanjing Agricultural University.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
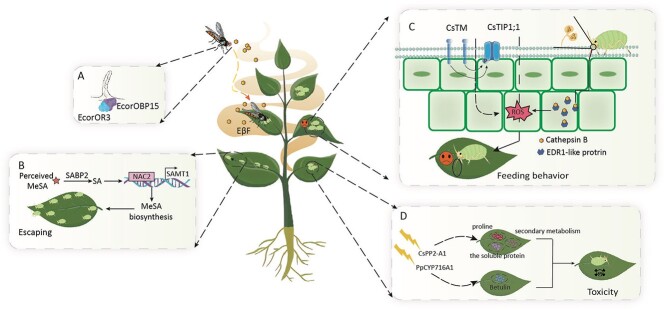
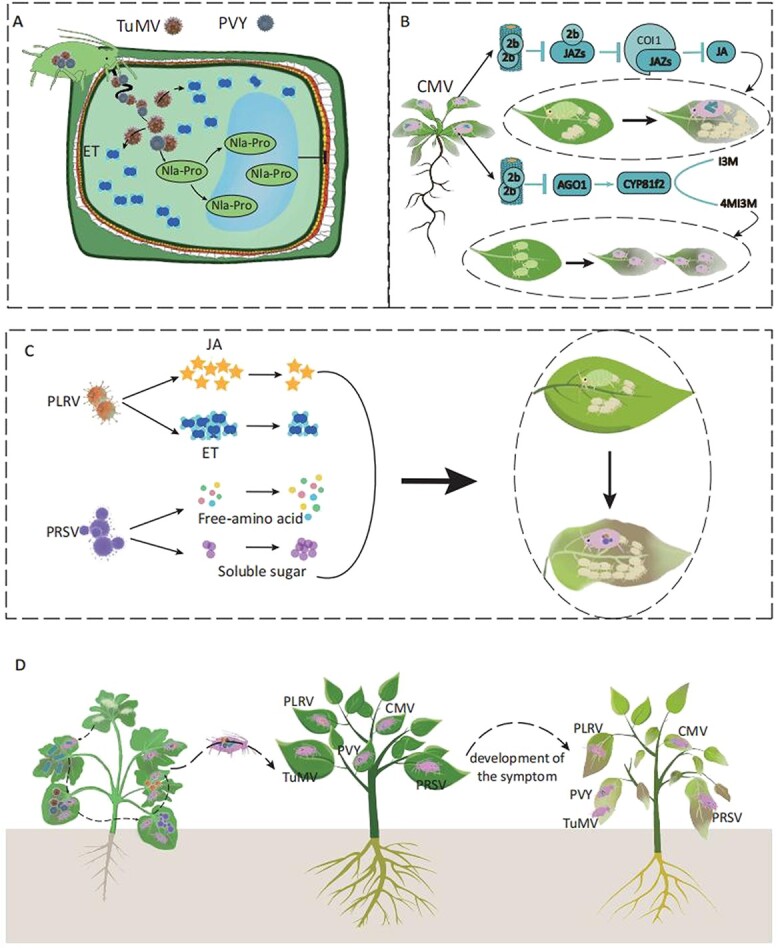
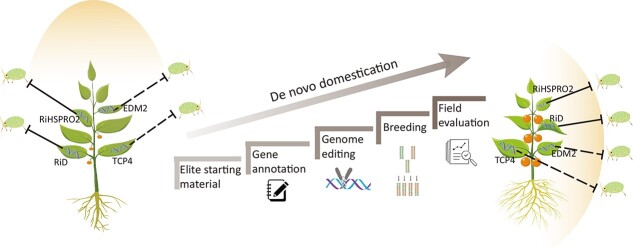
References
-
- Deutsch CA, Tewksbury JJ, Tigchelaar M. et al. Increase in crop losses to insect pests in a warming climate. Science. 2018;361:916–9 - PubMed
-
- Wang J, Li Y, Wang X. et al. Betulin, synthesized by Ppcyp716a1, is a key endogenous defensive metabolite of peach against aphids. J Agric Food Chem. 2022;70:12865–77 - PubMed
-
- De Kesel J, Conrath U, Flors V. et al. The induced resistance lexicon: do's and don'ts. Trends Plant Sci. 2021;26:685–91 - PubMed
LinkOut - more resources
Full Text Sources
