

This is a preprint.
RNA encodes physical information
- PMID: 39713325
- PMCID: PMC11661273
- DOI: 10.1101/2024.12.11.627970
RNA encodes physical information
Abstract
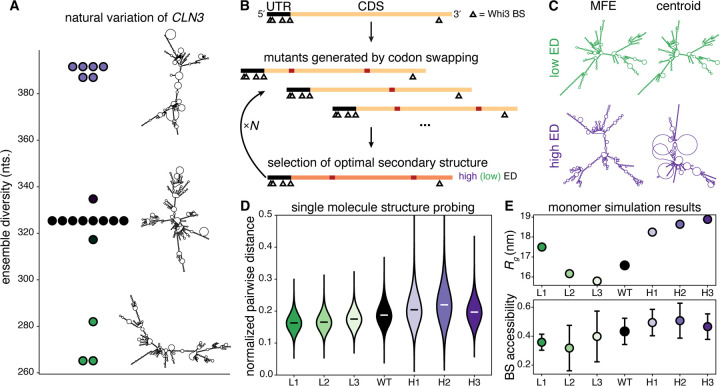
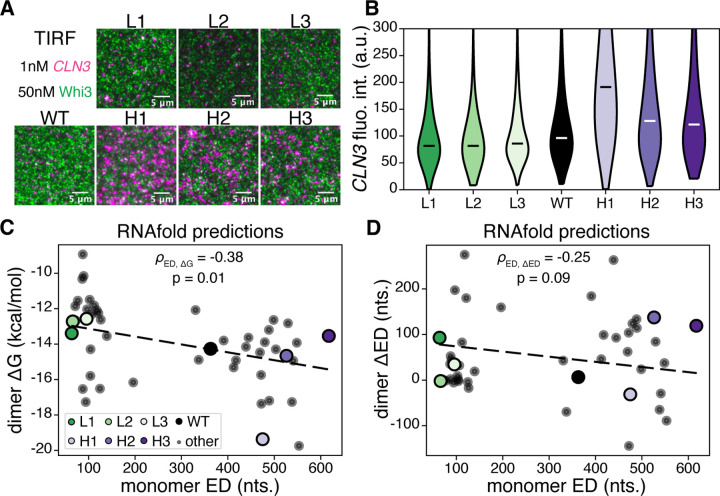
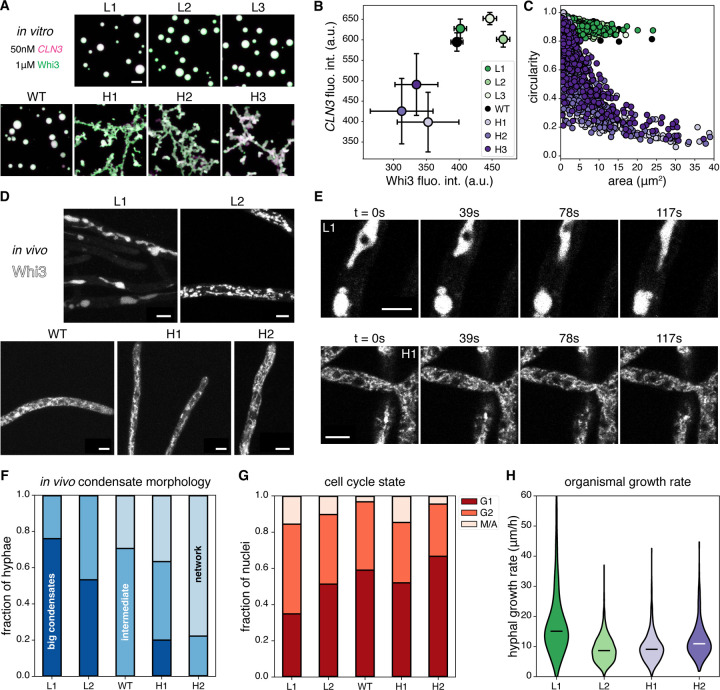
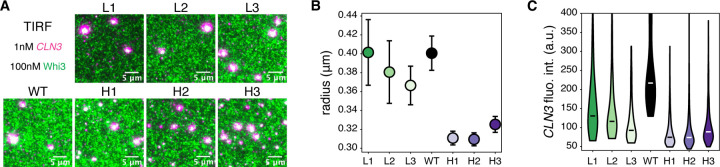
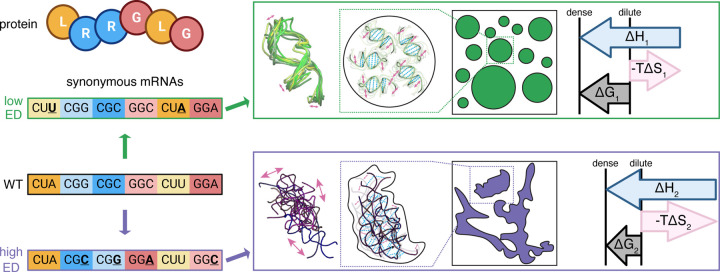
Most amino acids are encoded by multiple codons, making the genetic code degenerate. Synonymous mutations affect protein translation and folding, but their impact on RNA itself is often neglected. We developed a genetic algorithm that introduces synonymous mutations to control the diversity of structures sampled by an mRNA. The behavior of the designed mRNAs reveals a physical code layered in the genetic code. We find that mRNA conformational heterogeneity directs physical properties and functional outputs of RNA-protein complexes and biomolecular condensates. The role of structure and disorder of proteins in biomolecular condensates is well appreciated, but we find that RNA conformational heterogeneity is equally important. This feature of RNA enables both evolution and engineers to build cellular structures with specific material and responsive properties.
Keywords: RNA structure; biomolecular condensates; biopolymers; ensemble diversity; information theory; synonymous mutation.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources