

A novel human pluripotent stem cell gene activation system identifies IGFBP2 as a mediator in the production of haematopoietic progenitors in vitro
- PMID: 39714446
- PMCID: PMC11666236
- DOI: 10.7554/eLife.94884
A novel human pluripotent stem cell gene activation system identifies IGFBP2 as a mediator in the production of haematopoietic progenitors in vitro
Abstract
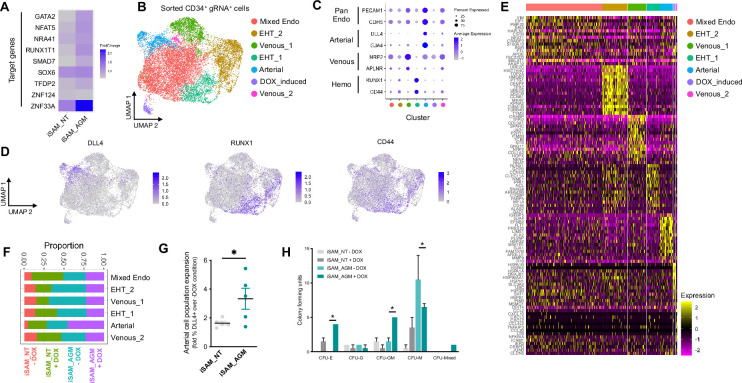
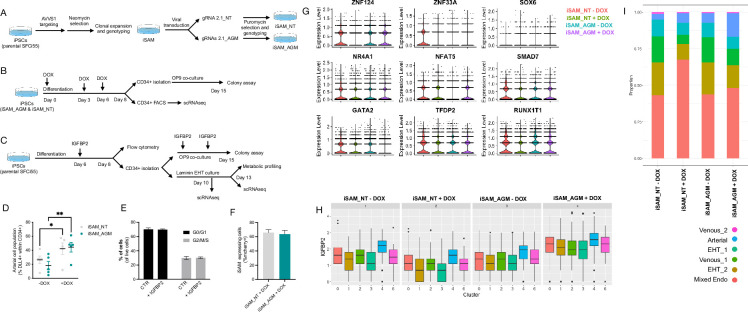
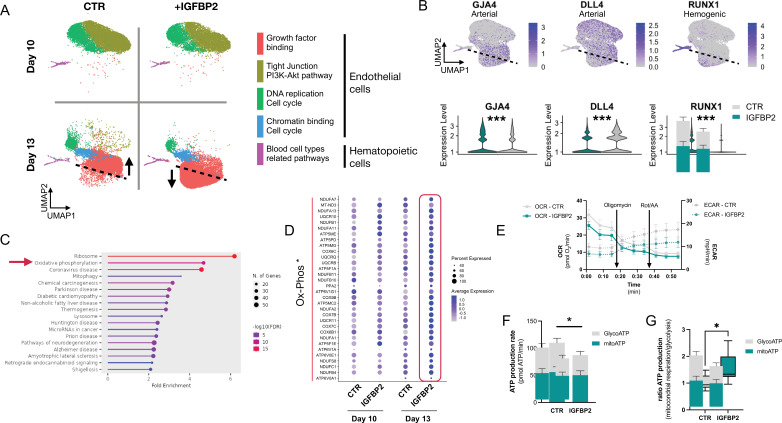
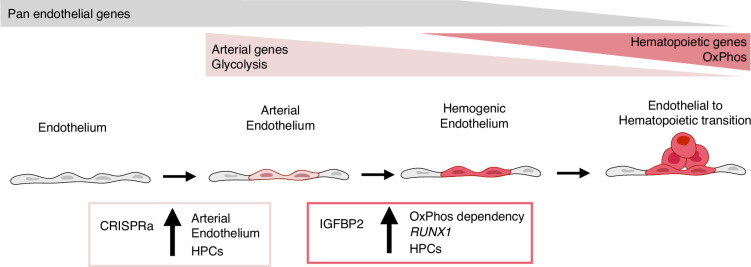
A major challenge in the stem cell biology field is the ability to produce fully functional cells from induced pluripotent stem cells (iPSCs) that are a valuable resource for cell therapy, drug screening, and disease modelling. Here, we developed a novel inducible CRISPR-mediated activation strategy (iCRISPRa) to drive the expression of multiple endogenous transcription factors (TFs) important for in vitro cell fate and differentiation of iPSCs to haematopoietic progenitor cells. This work has identified a key role for IGFBP2 in developing haematopoietic progenitors. We first identified nine candidate TFs that we predicted to be involved in blood cell emergence during development, then generated tagged gRNAs directed to the transcriptional start site of these TFs that could also be detected during single-cell RNA sequencing (scRNAseq). iCRISPRa activation of these endogenous TFs resulted in a significant expansion of arterial-fated endothelial cells expressing high levels of IGFBP2, and our analysis indicated that IGFBP2 is involved in the remodelling of metabolic activity during in vitro endothelial to haematopoietic transition. As well as providing fundamental new insights into the mechanisms of haematopoietic differentiation, the broader applicability of iCRISPRa provides a valuable tool for studying dynamic processes in development and for recapitulating abnormal phenotypes characterised by ectopic activation of specific endogenous gene expression in a wide range of systems.
Keywords: CRISPR activation; developmental biology; haematopoiesis; human; pluripotent stem cells; regenerative medicine; stem cells.
© 2024, Petazzi et al.
Conflict of interest statement
PP, TV, FL, HM, AM, HT, MS, NR, LF, PM, AF No competing interests declared
Figures







Update of
- doi: 10.1101/2024.01.14.575573
- doi: 10.7554/eLife.94884.1
- doi: 10.7554/eLife.94884.2
References
-
- afidanza CRISPRa. swh:1:rev:a65690d5082d7e9d08827e9d766b28430e4335c3Software Heriatge. 2022 https://archive.softwareheritage.org/swh:1:dir:a128c50858298df5c9e333b9a...
-
- Azzoni E, Frontera V, Anselmi G, Rode C, James C, Deltcheva EM, Demian AS, Brown J, Barone C, Patelli A, Harman JR, Nicholls M, Conway SJ, Morrissey E, Jacobsen SEW, Sparrow DB, Harris AL, Enver T, de Bruijn MFTR. The onset of circulation triggers a metabolic switch required for endothelial to hematopoietic transition. Cell Reports. 2021;37:110103. doi: 10.1016/j.celrep.2021.110103. - DOI - PMC - PubMed
-
- Böiers C, Carrelha J, Lutteropp M, Luc S, Green JCA, Azzoni E, Woll PS, Mead AJ, Hultquist A, Swiers G, Perdiguero EG, Macaulay IC, Melchiori L, Luis TC, Kharazi S, Bouriez-Jones T, Deng Q, Pontén A, Atkinson D, Jensen CT, Sitnicka E, Geissmann F, Godin I, Sandberg R, de Bruijn MFTR, Jacobsen SEW. Lymphomyeloid contribution of an immune-restricted progenitor emerging prior to definitive hematopoietic stem cells. Cell Stem Cell. 2013;13:535–548. doi: 10.1016/j.stem.2013.08.012. - DOI - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- S002219/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- EHA RAG 2021/European Hematology Association
- Research Global Award/American Society for Hematology
- Precision Medicine PhD scholarship/MRC_/Medical Research Council/United Kingdom
- Tissue Repair PhD studentship/College of Medicine and Veterinary Medicine, University of Edinburgh
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
