

Polyploids broadly generate novel haplotypes from trans-specific variation in Arabidopsis arenosa and Arabidopsis lyrata
- PMID: 39715277
- PMCID: PMC11706510
- DOI: 10.1371/journal.pgen.1011521
Polyploids broadly generate novel haplotypes from trans-specific variation in Arabidopsis arenosa and Arabidopsis lyrata
Abstract
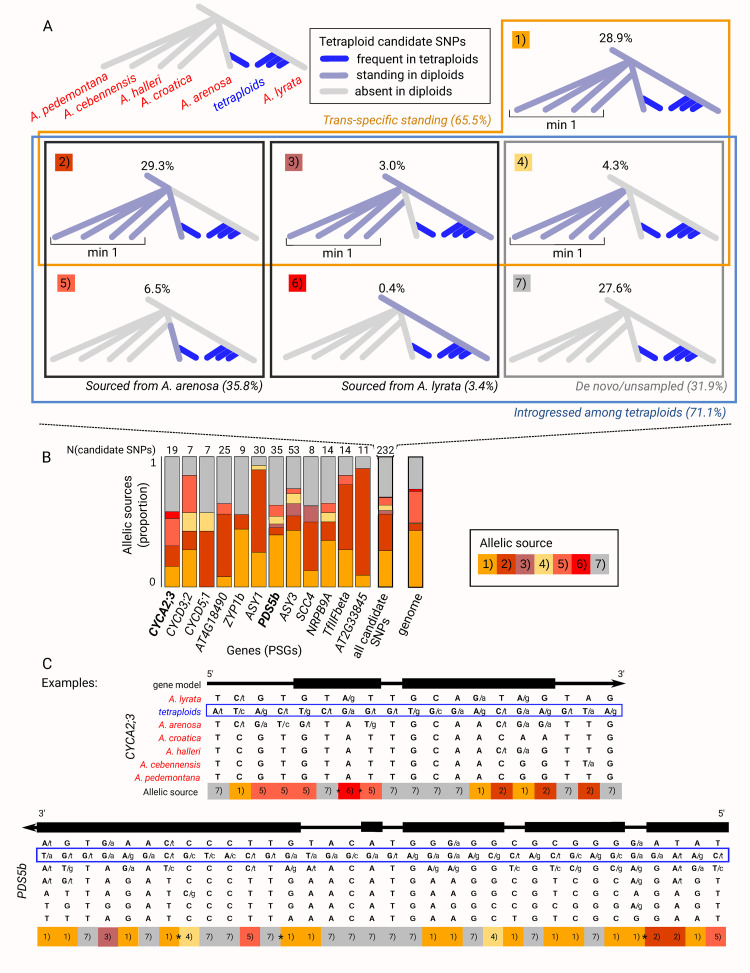
Polyploidy, the result of whole genome duplication (WGD), is widespread across the tree of life and is often associated with speciation and adaptability. It is thought that adaptation in autopolyploids (within-species polyploids) may be facilitated by increased access to genetic variation. This variation may be sourced from gene flow with sister diploids and new access to other tetraploid lineages, as well as from increased mutational targets provided by doubled DNA content. Here, we deconstruct in detail the origins of haplotypes displaying the strongest selection signals in established, successful autopolyploids, Arabidopsis lyrata and Arabidopsis arenosa. We see strong signatures of selection in 17 genes implied in meiosis, cell cycle, and transcription across all four autotetraploid lineages present in our expanded sampling of 983 sequenced genomes. Most prominent in our results is the finding that the tetraploid-characteristic haplotypes with the most robust signals of selection were completely absent in all diploid sisters. In contrast, the fine-scaled variant 'mosaics' in the tetraploids originated from highly diverse evolutionary sources. These include widespread novel reassortments of trans-specific polymorphism from diploids, new mutations, and tetraploid-specific inter-species hybridization-a pattern that is in line with the broad-scale acquisition and reshuffling of potentially adaptive variation in tetraploids.
Copyright: © 2024 Bohutínská et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Genetic adaptation associated with genome-doubling in autotetraploid Arabidopsis arenosa.PLoS Genet. 2012;8(12):e1003093. doi: 10.1371/journal.pgen.1003093. Epub 2012 Dec 20. PLoS Genet. 2012. PMID: 23284289 Free PMC article.
-
Single Geographic Origin of a Widespread Autotetraploid Arabidopsis arenosa Lineage Followed by Interploidy Admixture.Mol Biol Evol. 2015 Jun;32(6):1382-95. doi: 10.1093/molbev/msv089. Epub 2015 Apr 10. Mol Biol Evol. 2015. PMID: 25862142
-
Selection on meiosis genes in diploid and tetraploid Arabidopsis arenosa.Mol Biol Evol. 2015 Apr;32(4):944-55. doi: 10.1093/molbev/msu398. Epub 2014 Dec 26. Mol Biol Evol. 2015. PMID: 25543117 Free PMC article.
-
Polyploid Arabidopsis species originated around recent glaciation maxima.Curr Opin Plant Biol. 2018 Apr;42:8-15. doi: 10.1016/j.pbi.2018.01.005. Epub 2018 Feb 12. Curr Opin Plant Biol. 2018. PMID: 29448159 Review.
-
Adaptive introgression: how polyploidy reshapes gene flow landscapes.New Phytol. 2021 Apr;230(2):457-461. doi: 10.1111/nph.17204. Epub 2021 Feb 19. New Phytol. 2021. PMID: 33454987 Review.
Cited by
-
Whole-genome duplication increases genetic diversity and load in outcrossing Arabidopsis arenosa.Proc Natl Acad Sci U S A. 2025 Aug 5;122(31):e2501739122. doi: 10.1073/pnas.2501739122. Epub 2025 Jul 30. Proc Natl Acad Sci U S A. 2025. PMID: 40737318
-
Multiple Autopolyploid Arabidopsis lyrata Populations Stabilized by Long-Range Adaptive Introgression Across Eurasia.Mol Biol Evol. 2025 Jul 30;42(8):msaf153. doi: 10.1093/molbev/msaf153. Mol Biol Evol. 2025. PMID: 40701836 Free PMC article.
-
Rapid formation of stable autotetraploid rice from genome-doubled F1 hybrids of japonica-indica subspecies.Nat Plants. 2025 Apr;11(4):743-760. doi: 10.1038/s41477-025-01966-2. Epub 2025 Mar 31. Nat Plants. 2025. PMID: 40164786 Free PMC article.
-
Bridging micro and macroevolution: insights from chromosomal dynamics in plants.Front Plant Sci. 2025 Aug 22;16:1606450. doi: 10.3389/fpls.2025.1606450. eCollection 2025. Front Plant Sci. 2025. PMID: 40918955 Free PMC article. Review.
References
-
- Doyle JJ, Coate JE. Polyploidy, the nucleotype, and novelty: the impact of genome doubling on the biology of the cell. Int J Plant Sci. 2019;180(1): 1–52.
