

Leucine zipper-based immunomagnetic purification of CAR T cells displaying multiple receptors
- PMID: 39715901
- PMCID: PMC11917073
- DOI: 10.1038/s41551-024-01287-3
Leucine zipper-based immunomagnetic purification of CAR T cells displaying multiple receptors
Abstract
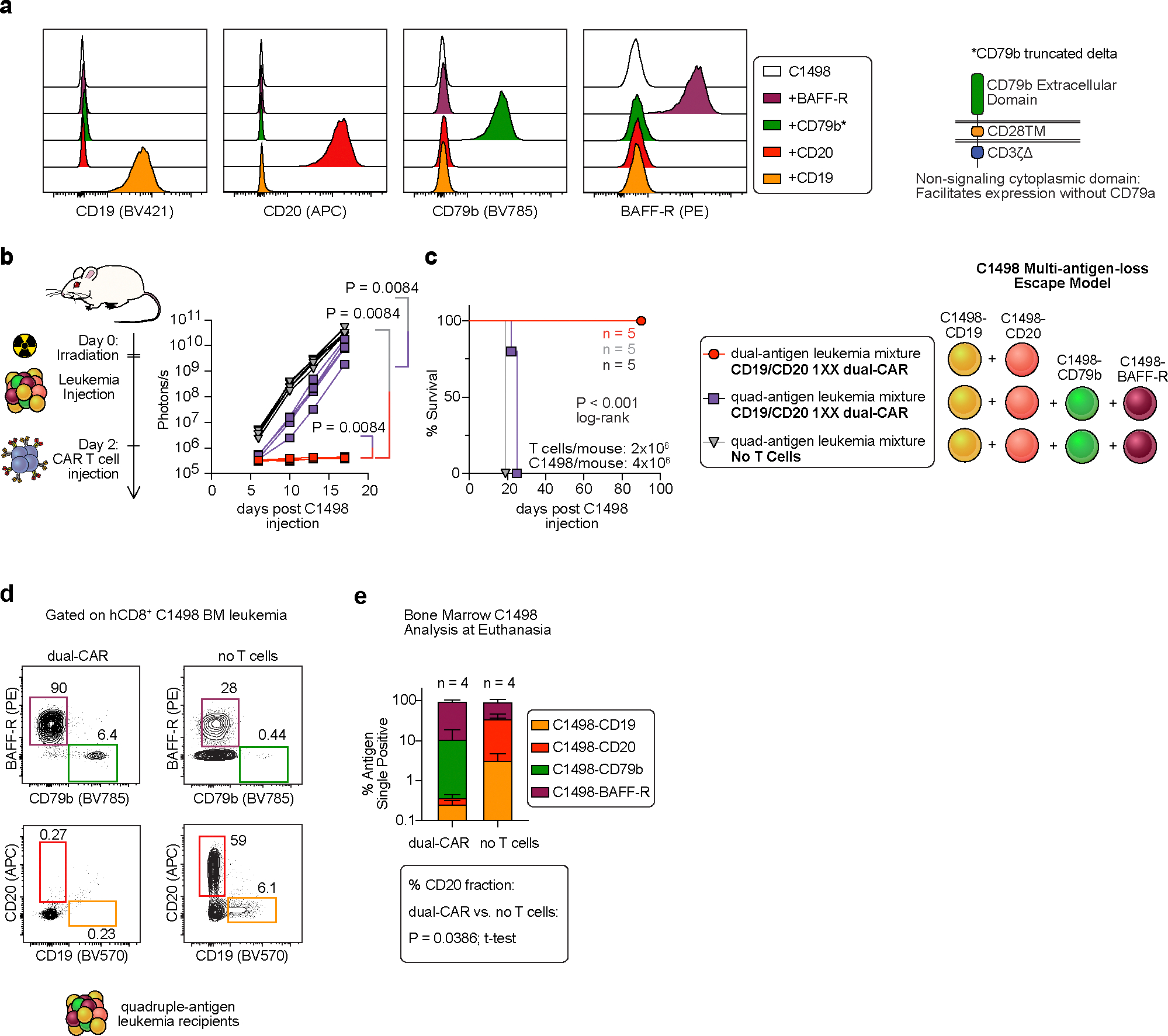
Resistance to chimaeric antigen receptor (CAR) T cell therapy develops through multiple mechanisms, most notably antigen loss and tumour-induced immune suppression. It has been suggested that T cells expressing multiple CARs may overcome the resistance of tumours and that T cells expressing receptors that switch inhibitory immune-checkpoint signals into costimulatory signals may enhance the activity of the T cells in the tumour microenvironment. However, engineering multiple features into a single T cell product is difficult because of the transgene-packaging constraints of current gene-delivery vectors. Here we describe a cell-sorting method that leverages leucine zippers for the selective single-step immunomagnetic purification of cells co-transduced with two vectors. Such 'Zip sorting' facilitated the generation of T cells simultaneously expressing up to four CARs and coexpressing up to three 'switch' receptors. In syngeneic mouse models, T cells with multiple CARs and multiple switch receptors eliminated antigenically heterogeneous populations of leukaemia cells coexpressing multiple inhibitory ligands. By combining diverse therapeutic strategies, Zip-sorted multi-CAR multi-switch-receptor T cells can overcome multiple mechanisms of CAR T cell resistance.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: S.E.J., L.J., and M.R.M.v.d.B. are co-inventors on patent applications related to this manuscript (‘Leucine zipper-based compositions and methods of use’, nos. US20210171601A1 (USA), EP3836944A4 (Europe), WO2020037178A1 (WIPO), CA3109630A1 (Canada) and CN112930186A (China); ‘Cell sorting systems and methods of use’, nos. US20210179686A1 (USA), EP3837287A4 (Europe), WO2020037181A2 (WIPO), CA3109635A1 (Canada) and CN112996819A (China)). A.P.B. has consulted for Bristol Myers Squibb and Cancer Study Group, LLC, and has received honoraria from OncLive. F.D.B. is a founder of Biocept and has intellectual property licensed to Novartis. J.U.P. reports research funding, intellectual property fees and travel reimbursement from Seres Therapeutics as well as consulting fees from Da Volterra, CSL Behring and MaaT Pharma. He serves on an Advisory board of, and holds equity in, Postbiotics Plus Research. He has filed intellectual property applications related to the microbiome. S.A.V. has received funding from Bristol-Meyers Squibb and has received consulting fees from Koch Disruptive Technologies and Generate Biomedicine. C.A.K. has previously filed intellectual property applications related to the FasDNR featured in this manuscript. C.A.K. is a scientific co-founder and holds equity in Affini-T Therapeutics. C.A.K. has previously consulted for or is on the scientific and/or clinical advisory boards of: Achilles Therapeutics, Affini-T Therapeutics, Aleta BioTherapeutics, Bellicum Pharmaceuticals, Bristol Myers Squibb, Catamaran Bio, Cell Design Labs, Decheng Capital, G1 Therapeutics, Klus Pharma, Obsidian Therapeutics, PACT Pharma, Roche/Genentech, Royalty Pharma and T-knife. M.R.M.v.d.B. has received research support and stock options from Seres Therapeutics and stock options from Notch Therapeutics and Pluto Therapeutics; has received royalties from Wolters Kluwer; has consulted, received honoraria from or participated in advisory boards for Seres Therapeutics, Vor Biopharma, Rheos Medicines, Frazier Healthcare Partners, Nektar Therapeutics, Notch Therapeutics, Ceramedix, Lygenesis, Pluto Therapeutics, GlaxoSmithKline, Da Volterra, Thymofox, Garuda, Novartis (spouse), Synthekine (spouse), Beigene (spouse), Kite (spouse); has intellectual property licensing with Seres Therapeutics and Juno Therapeutics; and holds a fiduciary role on the Foundation Board of DKMS (a non-profit organization). MSKCC has institutional financial interests relative to Seres Therapeutics.
Figures
















References
-
- Mikkilineni L & Kochenderfer JN CAR T cell therapies for patients with multiple myeloma. Nat. Rev. Clin. Oncol. 18, 71–84 (2021). - PubMed
-
- Berger TR & Maus MV Mechanisms of response and resistance to CAR T cell therapies. Curr. Opin. Immunol. 69, 56–64 (2021). - PubMed
-
- Majzner RG & Mackall CL Tumor Antigen Escape from CAR T-cell Therapy. Cancer Discov. 8, 1219–1226 (2018). - PubMed
-
- Labanieh L & Mackall CL CAR immune cells: design principles, resistance and the next generation. Nature 614, 635–648 (2023). - PubMed
MeSH terms
Substances
Grants and funding
- K08 CA237731/CA/NCI NIH HHS/United States
- R01-CA228308/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R01 HL147584/HL/NHLBI NIH HHS/United States
- R01 CA286507/CA/NCI NIH HHS/United States
- K08-CA252157/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R01-HL147584/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- P50 CA217694/CA/NCI NIH HHS/United States
- R01-CA228358/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R01-HL123340/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- K08 HL143189/HL/NHLBI NIH HHS/United States
- K08 CA252157/CA/NCI NIH HHS/United States
- K08 CA293271/CA/NCI NIH HHS/United States
- R35 CA284024/CA/NCI NIH HHS/United States
- R37 CA259177/CA/NCI NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
