

Similar deficiencies, different outcomes: succinate dehydrogenase loss in adrenal medulla vs. fibroblast cell culture models of paraganglioma
- PMID: 39716277
- PMCID: PMC11668036
- DOI: 10.1186/s40170-024-00369-9
Similar deficiencies, different outcomes: succinate dehydrogenase loss in adrenal medulla vs. fibroblast cell culture models of paraganglioma
Abstract
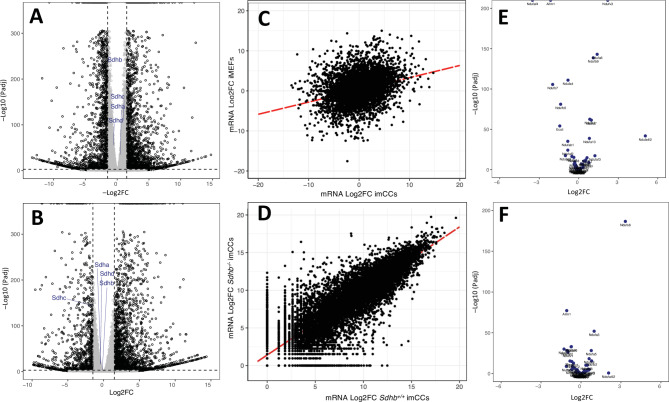
Heterozygosity for loss-of-function alleles of the genes encoding the four subunits of succinate dehydrogenase (SDHA, SDHB, SDHC, SDHD), as well as the SDHAF2 assembly factor predispose affected individuals to pheochromocytoma and paraganglioma (PPGL), two rare neuroendocrine tumors that arise from neural crest-derived paraganglia. Tumorigenesis results from loss of the remaining functional SDHx gene copy, leading to a cell with no functional SDH and a defective tricarboxylic acid (TCA) cycle. It is believed that the subsequent accumulation of succinate competitively inhibits multiple dioxygenase enzymes that normally suppress hypoxic signaling and demethylate histones and DNA, ultimately leading to increased expression of genes involved in angiogenesis and cell proliferation. Why SDH loss is selectively tumorigenic in neuroendocrine cells remains poorly understood. In the absence of SDH-loss tumor-derived cell models, the cellular burden of SDH loss and succinate accumulation have been investigated through conditional knockouts of SDH subunits in pre-existing murine or human cell lines with varying degrees of clinical relevance. Here we characterize two available murine SDH-loss cell lines, immortalized adrenally-derived premature chromaffin cells vs. immortalized fibroblasts, at a level of detail beyond that currently reported in the literature and with the intention of laying the foundation for future investigations into adaptive pathways and vulnerabilities in SDH-loss cells. We report different mechanistic and phenotypic manifestations of SDH subunit loss in the presented cellular contexts. These findings highlight similarities and differences in the cellular response to SDH loss between the two cell models. We show that adrenally-derived cells display more severe morphological cellular and mitochondrial alterations, yet are unique in preserving residual Complex I function, perhaps allowing them to better tolerate SDH loss, thus making them a closer model to SDH-loss PPGL relative to fibroblasts.(281 words).
Keywords: Complex I; Hypoxia; Paraganglioma; Pheochromocytoma; Succinate dehydrogenase; Tricarboxylic acid cycle.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests. Data deposition: RNA sequencing data have been deposited in the Gene Expression Omnibus (GEO) database as GSE263778.
Figures




Similar articles
-
Distinct proteomic and acylproteomic adaptations to succinate dehydrogenase loss in two cell contexts.bioRxiv [Preprint]. 2025 Aug 2:2025.08.01.668168. doi: 10.1101/2025.08.01.668168. bioRxiv. 2025. PMID: 40766370 Free PMC article. Preprint.
-
Metabolic Effects of Succinate Dehydrogenase Loss in Cancer.J Cell Physiol. 2025 Jul;240(7):e70066. doi: 10.1002/jcp.70066. J Cell Physiol. 2025. PMID: 40685935 Review.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
Update on Tumor Surveillance for Children with Hereditary Pheochromocytoma/Paraganglioma Syndromes.Clin Cancer Res. 2025 Aug 14;31(16):3368-3376. doi: 10.1158/1078-0432.CCR-24-4354. Clin Cancer Res. 2025. PMID: 40549645 Review.
Cited by
-
Distinct proteomic and acylproteomic adaptations to succinate dehydrogenase loss in two cell contexts.bioRxiv [Preprint]. 2025 Aug 2:2025.08.01.668168. doi: 10.1101/2025.08.01.668168. bioRxiv. 2025. PMID: 40766370 Free PMC article. Preprint.
-
Approaches to reduce succinate accumulation by restoration of succinate dehydrogenase activity in cultured adrenal cells.J Cell Sci. 2025 May 1;138(9):jcs263925. doi: 10.1242/jcs.263925. Epub 2025 May 12. J Cell Sci. 2025. PMID: 40235395 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
