

Ubiquitin-mediated recruitment of the ATG9A-ATG2 lipid transfer complex drives clearance of phosphorylated p62 aggregates
- PMID: 39718773
- PMCID: PMC11809316
- DOI: 10.1091/mbc.E24-03-0101
Ubiquitin-mediated recruitment of the ATG9A-ATG2 lipid transfer complex drives clearance of phosphorylated p62 aggregates
Abstract
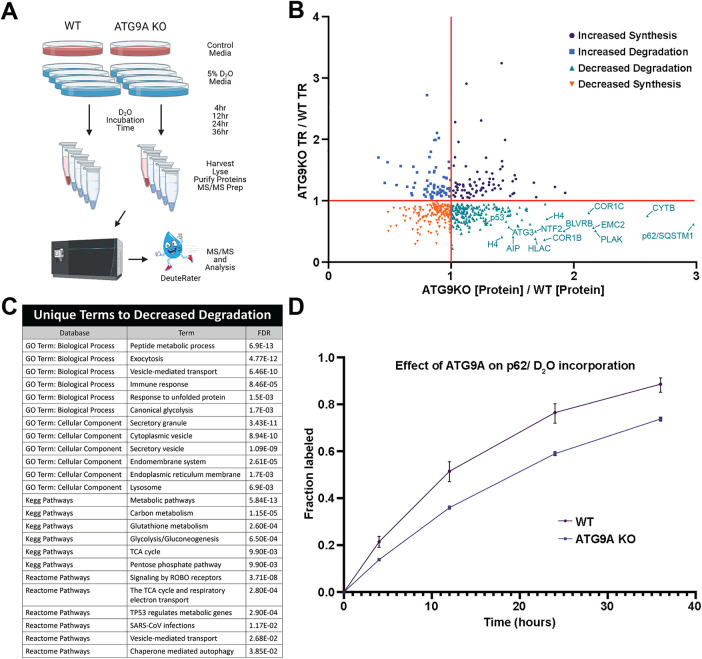
Autophagy is an essential cellular recycling process that maintains protein and organelle homeostasis. ATG9A vesicle recruitment is a critical early step in autophagy to initiate autophagosome biogenesis. The mechanisms of ATG9A vesicle recruitment are best understood in the context of starvation-induced nonselective autophagy, whereas less is known about the signals driving ATG9A vesicle recruitment to autophagy initiation sites in the absence of nutrient stress. Here we demonstrate that loss of ATG9A, or the lipid transfer protein ATG2, leads to the accumulation of phosphorylated p62 aggregates in nutrient replete conditions. Furthermore, we show that p62 degradation requires the lipid scramblase activity of ATG9A. Last, we present evidence that polyubiquitin is an essential signal that recruits ATG9A and mediates autophagy foci assembly in nutrient replete cells. Together, our data support a ubiquitin-driven model of ATG9A recruitment and autophagosome formation during basal autophagy.
Conflict of interest statement
Conflict of interest: The authors declare no financial conflicts of interest.
Figures




References
-
- Adriaenssens E, Ferrari L, Martens S (2022). Orchestration of selective autophagy by cargo receptors. Curr Biol 32, R1357–R1371. - PubMed
-
- Agudo-Canalejo J, Schultz SW, Chino H, Migliano SM, Saito C, Koyama-Honda I, Stenmark H, Brech A, May AI, Mizushima N, Knorr RL (2021). Wetting regulates autophagy of phase-separated compartments and the cytosol. Nature 591, 142–146. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
