

The cytokinin efflux transporter ABCC4 participates in Arabidopsis root system development
- PMID: 39719052
- PMCID: PMC11668331
- DOI: 10.1093/plphys/kiae628
The cytokinin efflux transporter ABCC4 participates in Arabidopsis root system development
Abstract
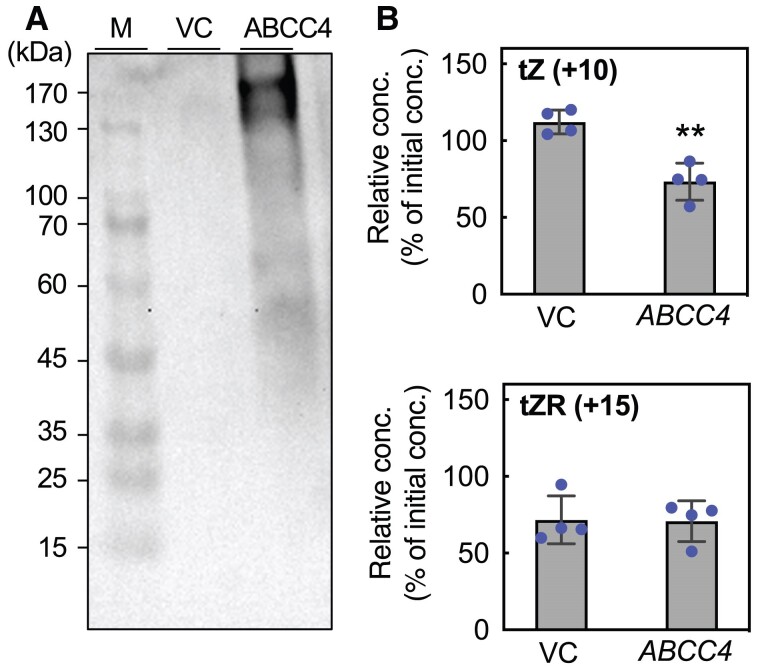
The directional and sequential flow of cytokinin in plants is organized by a complex network of transporters. Genes involved in several aspects of cytokinin transport have been characterized; however, much of the elaborate system remains elusive. In this study, we used a transient expression system in tobacco (Nicotiana benthamiana) leaves to screen Arabidopsis (Arabidopsis thaliana) transporter genes and isolated ATP-BINDING CASSETTE TRANSPORTER C4 (ABCC4). Validation through drug-induced expression in Arabidopsis and heterologous expression in budding yeast revealed that ABCC4 effluxes the active form of cytokinins. During the seedling stage, ABCC4 was highly expressed in roots, and its expression was upregulated in response to cytokinin application. Loss-of-function mutants of ABCC4 displayed enhanced primary root elongation, similar to mutants impaired in cytokinin biosynthesis or signaling, that was suppressed by exogenous trans-zeatin treatment. In contrast, overexpression of the gene led to suppression of root elongation. These results suggest that ABCC4 plays a role in the efflux of active cytokinin, thereby contributing to root growth regulation. Additionally, cytokinin-dependent enlargement of stomatal aperture was impaired in the loss-of-function and overexpression lines. Our findings contribute to unraveling the many complexities of cytokinin flow and enhance our understanding of the regulatory mechanisms underlying root system development and stomatal opening in plants.
© The Author(s) 2024. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures






References
-
- Ando E, Ohnishi M, Wang Y, Matsushita T, Watanabe A, Hayashi Y, Fujii M, Ma JF, Inoue S-I, Kinoshita T. TWIN SISTER OF FT, GIGANTEA, and CONSTANS have a positive but indirect effect on blue light-induced stomatal opening in Arabidopsis. Plant Physiol. 2013:162(3):1529–1538. 10.1104/pp.113.217984 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
